Assessment of Thyroglobulin Expression in Reproductive Organs at Different Stages of Mouse Estrous Cycle
-
Moravvej, Ali
-
Department of Immunology, Faculty of Medicine, Iran University of Medical Sciences , Tehran, Iran
-
Jeddi-Tehrani, Mahmood
-
Monoclonal Antibody Research Center, Avicenna Research Institute, ACECR , Tehran, Iran
-
Salek Moghaddam, Ali Reza
-
Department of Immunology, Faculty of Medicine, Iran University of Medical Sciences , Tehran, Iran
-
Dokouhaki, Pouneh
-
Reproductive Biotechnology Research Center, Avicenna Research Institute, ACECR , Tehran, Iran
-
Shekarabi, Mahdi
-
Department of Immunology, Faculty of Medicine, Iran University of Medical Sciences , Tehran, Iran
-
Ghods, Roya
-
Monoclonal Antibody Research Center, Avicenna Research Institute, ACECR , Tehran, Iran
-
Shahbazi, Mahdi
-
Department of Immunology, Faculty of Medicine, Iran University of Medical Sciences , Tehran, Iran
-
Ghasemi, Jamileh
-
Reproductive Biotechnology Research Center, Avicenna Research Institute, ACECR , Tehran, Iran
-
Danesh, Parivash
-
Department of Immunology, Faculty of Medicine, Iran University of Medical Sciences , Tehran, Iran
-
Mahmoudi, Ahmad Reza
-
Department of Immunology, Faculty of Medicine, Iran University of Medical Sciences , Tehran, Iran
-
 Zarnani, Amir-Hassan
Ph.D., Reproductive Biotechnology Research Center, Avicenna Research Institute, ACECR, Tehran, Iran, P.O. Box: 19615-1177, Tel: +98 21 22432020, Fax: +98 21 22432021, E-mail: zarnani25@yahoo.com
Zarnani, Amir-Hassan
Ph.D., Reproductive Biotechnology Research Center, Avicenna Research Institute, ACECR, Tehran, Iran, P.O. Box: 19615-1177, Tel: +98 21 22432020, Fax: +98 21 22432021, E-mail: zarnani25@yahoo.com
-
Reproductive Biotechnology Research Center, Avicenna Research Institute, ACECR , Tehran, Iran
-
Immunology Research Center, Faculty of Medicine, Iran University of Medical Sciences , Tehran, Iran
Abstract: Prevalence of abortion is higher in women with autoimmune thyroid disease. In the majority of cases, however, no abnormality of thyroid function is detected despite the high levels of antithyroid antibodies. The direct influence of such harmful autoantibodies in female reproductive organs may serve a role in pregnancy loss. In this study, expression of thyroglobulin in the reproductive tissues of cycling mice has been evaluated. Stages of estrous cycle were determined by cellular morphology and ratio of epithelial cells to leukocytes in vaginal smear of Balb/C mice. At each phase, the mice were sacrificed and their uterus, ovary and fallopian tubes were removed. Expression of thyroglobulin-specific transcript in endometrium was investigated by two sets of primers using reverse transcriptase-polymerase chain reaction (RT-PCR). In addition, expression of thyroglobulin in reproductive tissues was assessed by immunohistochemistry and dot blot analysis. The results showed that thyroglobulin mRNA is not expressed in endometrial tissue of Balb/C mice at any stage of estrous cycle. Immunohistochemical analysis also confirmed that thyroglobulin or its cross reactive-antigens are not expressed at the protein level in the female reproductive organs. The results showed that thyroglobulin was not expressed in the reproductive organs of female mice. It is plausible that antithyroglobulin antibodies could interact with newly-generated antigens during placentation and pregnancy.
Introduction :
Abortion is the most prevalent compli-cation of pregnancy. In more than half of the cases, the underlying cause of abortion is unknown. However, a variety of factors such as genetic abnormalities, structural problems, autoimmune diseases and infections have been known to cause abortion (1,2). Among the autoimmune diseases implicated to cause abortion and infertility, thyroid autoimmunity is thought to be the most prominent one (3-7). Most patients with thyroid autoimmunity have high serum levels of anti thyroglobulin (TG) and/or antithyroid peroxidase (TPO) antibodies. One mechanism by which these antibodies can cause infertility, abortion or congenital abnormalities in the fetus, is by direct binding to thyroid proteins vital for thyroid hormone production with consequent reduction in serum levels of thyroid hormones (8-10). In the majority of cases, however, no abnormality of thyroid function is detected despite high levels of antithyroid antibodies. It has been shown that recurrent abortion is still 2 to 4 times more frequent in these patients compared to normal women (3,4,11). Moreover, infertile women with high levels of peroxidase autoantibodies tend to have more abortions after receiving IVF treatment for their infertility (7,12). Indeed, maternal thyroid peroxidase antibodies during pregnancy are associated with impaired child development (13). Although there are some speculations as to how thyroid autoantibodies are linked to abortion, there is no direct evidence to address the issue. There are three main hypo-theses for this phenomenon:
- Antithyroid antibodies bind directly to their cognate antigens in the thyroid gland and interfere with production of thyroid hormones and hence reduce serum levels of the hormones;
- Antithyroid antibodies are among auto- antibodies that are produced during a general-ized autoimmune disease with disrupted immunological balance. In fact, the presence of antithyroid antibodies can be translated as the presence of abnormal immune reactions that lead to abortion;
- Antithyroid antibodies cross react with unknown antigenic epitope(s) in the reproduc-tive tissue and lead to local damage and abortion (4, 7).
Despite many efforts, there is still not enough evidence to confirm or refute any one of these hypotheses. In the present study, we aimed to investigate the expression of thyro-globulin gene in murine endometrium in order to evaluate the third possibility.
Materials and Methods :
Animals
In this study, white New Zealand rabbits and Balb/C mice aged 8-12 weeks were used. All procedures were conducted according to the guidelines of the animal care and ethics committee of Avicenna Research Institute. Determination of estrous cycle
Wet vaginal smear was obtained by intra vaginal injection of Phosphate Buffered Saline (PBS). Stages of estrous cycle were de-termined by cellular morphology and ratio of epithelial cells to leukocytes in each slide. Five mice were studied in each four stages of estrous cycle: proestrous, estrous, metestrous and diestrous. RNA extraction and complementary DNA (cDNA) synthesis
Following determination of estrous cycle, mice were sacrificed and their uterine horns were excised. Left uterine horn was washed completely with PBS and cut open from anti- mesometrial axis. Exposed endometrial tissue was then scraped with scalpel. Endometrial tissue was rapidly homogenized using pellet pestle (Sigma, USA) and dissolved in RNA-Bee (Biosite, Sweden).
As our positive control for thyroglobulin expression, thyroid gland of one mouse was excised and homogenized similarly in RNA-Bee solution. Total RNA from tissue homo-genates was then extracted according to standard protocols. Briefly, after chloroform was added to and mixed with RNA-Bee solution, total RNA of tissue was collected from liquid supernatant and precipitated by 2 steps alcohol precipitation. Extracted RNA was dissolved in 20-50 µl of sterile water and stored in -20ºC for further analysis.
Ten micro liters of total RNA from each tissue were heated to 65ºC and immediately cooled on ice. cDNA Mix including 5x buffer, 2 mM dNTP mix (Roche, Germany), 2 mM Random hexamer (Cybergene, Sweden) and 20 u/µl RT M-MuLV in a final volume of 10 µl distilled water was added to the RNA. The mixture was then incubated in 42ºC for 60 minutes. The cDNA was then kept at -20ºC.
Reverse transcriptase polymerase chain reaction (RT-PCR)
For each specimen, 3 PCR reactions were set up using 2 sets of primers for TG and 1 set for housekeeping gene GAPDH (Glycer al-dehyde 3-phosphate dehydrogenase). Each PCR reaction contained 10x PCR buffer (Roche, Germany), 2.5 mM MgCl2, 0.4 mM dNTP mix and 0.04 u/µl Taq DNA poly-merase (all from Roche, Germany) and 0.4 µM of each primer set. One microliter of cDNA mixture was added to each reaction and incubated in thermocycler with the following thermal profile: 94ºC for 2 minutes for initial denaturation, 30 cycles of 94ºC, 60ºC and 72ºC each for 30 seconds and final extension in 72ºC for 6 minutes. Sequence of the primers used is demonstrated in Table 1.
First and second sets of primers were designated to amplify 207 bp (TG1) and 510 bp (TG2) fragments of TG mRNA, respectively. The third set of primers amplified a 309 bp segment of GAPDH gene as internal control. PCR products were elec-trophoresed on 1% agarose gel and the amplified bands were visualized and docu-mented by UV transilluminator (UVP, USA).
Generation of polyclonal antiTG antibody
White New Zealand rabbits were injected IM with bovine TG (Sigma, Sweden) plus freund adjuvant every 2 weeks. Before and after each injection, blood samples were collected and titers of anti TG antibody were assessed by indirect ELISA. Briefly, TG was coated on wells and after blocking, rabbit serum was added to them in serial dilutions. Following incubation and washing, HRP con-jugated sheep antirabbit antibody (Avicenna Research Institute, Iran) was added to the wells.
Finally, optical density at 450 nm was measured after addition of horseradish per-oxidase (HRP) substrate, TMB (3, 3', 5, 5'-tetra methyl benzidine).
Following the 5th injection, massive bleeding was performed and the antiTG antibody was purified over TG affinity chro-matography column. Purity of the resultant antibody was assessed by electrophoresis on 10% polyacrylamide gel and its reactivity was confirmed by immunohistochemistry of mouse th
Result :
Amplification of TG gene by RT-PCR
As a housekeeping gene, GAPDH was used as an internal control for mRNA extrac-tion and cDNA synthesis. As expected, 309 bp band of amplified GAPDH mRNA was found in all endometrial and thyroid samples. On the contrary, neither 207 bp nor 510 bp bands corresponding to the amplified segments of TG mRNA with TG1 & TG2 primer sets, respectively, were detected in any endometrial samples but they were both strongly positive in thyroid tissue. Therefore, we concluded that TG gene is not expressed
Generation and characterization of antiTG antibody
Despite the fact that we used bovine TG for immunization, the resulting polyclonal antibody had excellent reactivity with mouse TG because of high molecular homology between species. Based on ELISA assay, high titers of antiTG antibody were reached after the third round of injection.
Polyacrylamide gel electrophoresis of the purified antibody showed a dominant band of 150 kD molecular weight which corresponded to IgG isotype. In order to confirm the reactivity of the purified antibody, we used IHC staining on both frozen and paraffin embedded sections of thyroid which showed specific binding in both methods (Figure 2).
IHC staining of reproductive organs
In order to study the expression of thyroglobulin at the protein level in female reproductive organs, we performed IHC staining of endometrium, ovary and fallopian tubes at different stages of estrous cycle using our homemade antiTG antibody.
As shown in Figure 3, none of the tissues of reproductive organs had detectable levels of TG, while sections of thyroid were all strongly positive for expression of TG. IHC staining confirmed our RT-PCR results and proved that TG is expressed at neither RNA nor protein level in any reproductive organs of cycling Balb/C mice.
Discussion :
Autoimmune thyroid disease is one of the disorders that has a known role in recurrent abortion and infertility (3). Even in the absence of clinical manifestations of thyroid dysfunc-tion, many cases have high titers of anti-thyroid antibodies in their serum. Although dysfunctional production of thyroid hormones can directly cause abortion, it does not fully explain the correlation between abortion and thyroid autoimmunity. It has been well docu-mented that cases with completely normal serum levels of thyroid hormones, but with high levels of antiTG antibodies have increased risk of abortion by at least two fold compared to normal women (14).
This increased risk could as well be explained by direct binding of thyroid auto-antibodies to reproductive organs or to fetal tissues (7,15). In order to test this hypothesis, we studied the expression of TG at both mRNA and protein levels in all reproductive organs of female Balb/C mice. Since many proteins are modulated in reproductive tract under the influence of hormonal changes (16), we also tested the possibility of expression of TG gene in different stages of estrous cycle. It was clearly demonstrated that TG gene is expressed in endometrium during none of the four stages of estrous cycle. Also, we showed that no TG or TG-cross reactive antigen exists in either endometrium, ovary or fallopian tubes. Although endometrial cells of non-pregnant uterus are the same cells that comprise pregnant one, their pattern of gene expression undergoes dramatic changes during pregnancy (16). It could then be plausible that some genes, such as TG, have temporary expression only during pregnancy and therefore, study of their expression in non-pregnant state might not completely reflect the whole profile of these cells. In line with this hypothesis, it has been shown that pregnancy rate of women with antithyroid antibody was the same as normal controls but they experienced high rates of abortion afterwards (7,17). It could be implicated that TG or cross reactive antigen(s) are expressed at the interface of mother and fetus and this results in their interaction with maternal anti- TG antibodies. Subsequent inflammation at the site of pregnancy then ends up in abortion. Other studies show that active immunization of pregnant mice with TG resulted in lower weight of fetus and placenta. In the same study, it was shown that maternal antiTG antibodies were able to cross placenta and enter amniotic fluid and bind directly to syncytio- and cytotrophoblasts but not to fetus (17). Transient expression of TG in decidua, placenta and fetal tissues during pregnancy is the subject of the future studies.

Figure 1. Amplification of TG gene in endometrium of cycling mice by RT-PCR Endometrium of cycling mice was pealed away during each stage of estrous cycle and expression of TG gene was assessed by two sets of primers (TG1 & TG2). GAPDH amplification was used as internal control for RNA extraction and cDNA synthesis. 309bp band of amplified GAPDH mRNA was found in all endometrial and thyroid samples. On the contrary, 207bp band corresponding to the amplified segments of TG1 (a) or 510bp band corresponding to the amplified segments of TG2 (b) were not detected in any endometrial samples but it was strongly detected in thyroid tissue.
|
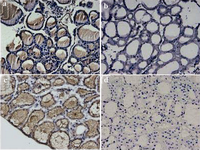
Figure 2. Immunostaining of mouse thyroid with polyclonal rabbit antiTG antibody To confirm reactivity of homemade antiTG antibody, immunohistochemistry staining of frozen (a) and paraffin embedded (c) sections of mouse thyroid was performed. In both methods the antibody showed strong immonoreactivity. b and d: Negative control slides of frozen and paraffin embedded sections, respectively. (Magnification: 200 �).
|
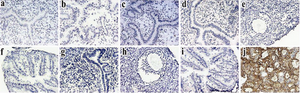
Figure 3. Immunohistochemical analysis of reproductive organs of cycling mice with antiTG antibody Uterus, ovary and fallopian tubes of cycling Balb/C mice were tested by immunohistochemistry for expression of TG. TG was not expressed in any of the tissues at all stages of estrous cycle. a-d: endometrium in proestrous, estrus, metestrous and diestrous, respectively. e and f: representative of ovary and fallopian tube. g-i: negative controls of endometrium, ovary and fallopian tubes. j: thyroid as positive control.
|

Tabel 1
|
|