Antifungal Indole and Pyrrolidine-2,4-Dione Derivative Peptidomimetic Lead Design Based on In Silico Study of Bioactive Peptide Families
-
Moradi, Shoeib
-
Drug Design and Bioinformatics Unit, Medical Biotechnology Department, Biotechnology Research Center, Pasteur Institute of Iran, Tehran, Iran
-
Azerang, Parisa
-
Drug Design and Bioinformatics Unit, Medical Biotechnology Department, Biotechnology Research Center, Pasteur Institute of Iran, Tehran, Iran
-
Khalaj, Vahid
-
Fungal Biotechnology Group, Medical Biotechnology Department, Biotechnology Research Center, Pasteur Institute of Iran, Tehran, Iran
-
 Sardari, Soroush
Drug Design and Bioinformatics Unit, Medical Biotechnology Department, Biotechnology Research Center, Pasteur Institute, Tehran, Iran, +98 21 66480780; Email: sardari@pasteur.ac.ir
Sardari, Soroush
Drug Design and Bioinformatics Unit, Medical Biotechnology Department, Biotechnology Research Center, Pasteur Institute, Tehran, Iran, +98 21 66480780; Email: sardari@pasteur.ac.ir
-
Drug Design and Bioinformatics Unit, Medical Biotechnology Department, Biotechnology Research Center, Pasteur Institute of Iran, Tehran, Iran
Abstract: Background: The rise of opportunistic fungal infections highlights the need for development of new antimicrobial agents. Antimicrobial Peptides (AMPs) and Antifungal Peptides (AFPs) are among the agents with minimal resistance being developed against them, therefore they can be used as structural templates for design of new antimicrobial agents.
Methods: In the present study four antifungal peptidomimetic structures named C1 to C4 were designed based on plant defensin of Pisum sativum. Minimum inhibitory concentrations (MICs) for these structures were deter-mined against Aspergillus niger N402, Candida albicans ATCC 10231, and Saccharomyces cerevisiae PTCC 5052.
Results: C1 and C2 showed more potent antifungal activity against these fun-gal strains compared to C3 and C4. The structure C2 demonstrated a potent antifungal activity among them and could be used as a template for future study on antifungal peptidomemetics design. Sequences alignments led to identifying antifungal decapeptide (KTCENLADTY) named KTC-Y, which its MIC was determined on fungal protoplast showing 25 (μg/ml) against Asper-gillus fumigatus Af293.
Conclusion: The present approach to reach the antifungal molecules seems to be a powerful approach in design of bioactive agents based on AMP mimetic identification.
Introduction :
In the past 10 years, the incidence of fungal infections has increased obviously 1. The emerging multiple-drug resistance has led to induce research for the detection of novel drugs through various mechanisms. AMPs and AFPs are evolutionarily preserved modules of the innate immune response and are established along the entire classes of life 2. These peptides are effective, broad spectrum antibiotics which reveal potential as new therapeutic agents 3. These AMPs are distributed in various sources ranging from primary eukaryotes to mammalians containing normally less than 60 amino acids long, having cationic amino acid residues and an amphiphilic structure bound to the membrane (α-helical and/or β-sheet) 4,5. The application of these antifungal peptides as drugs is not easy due to their poor oral and tissue absorption, rapid proteolytic cleavage and weak half-life or stability. Since the majority of proteins and small peptides are simply proteolyzed, quickly excreted and poorly bioavailable, attempts have been made to determine the ways of replacing peptide portions. These structures are termed as peptidomimetics which are to mimic peptides in the anticipation of attaining more bioavailable units 6. Their artificial backbone is resistant to proteases as well. Peptidomimetics are one set of probes utilized in the shift pathway of tiny molecule drug design.
Recently, significant progress has been made in the use of Computer-Aided Molecular Design (CAMD) of novel molecules with desired properties. These methods typically rely on two stages: the first stage is forward modelling 7, through which Quantitative Structure Activity Relationship (QSAR) process is accomplished by application of non-linear modelling procedures such as Artificial Neural Networks (ANN). As example of this strategy, in our group 8, over 100 antifungal peptides were analyzed by artificial neural networks and observed that most important physicochemical parameters affecting bioactivity of antifungal peptides are Log P and relative amphipaticity.
In the present study, 4 antifungal peptidomimetics structures were designed based on antifungal peptide structures, using bioinformatics and peptidomimetics strategy. The designed compounds were prepared and tested for antimicrobial activity.
Materials and Methods :
Data collection: For this study, more than 60 antifungal peptides were selected. Their amino acids sequences were used as input data for sequences alignments in order to find out the shared amino acids sequences among them.
Sequences alignments: T-Coffee v5.13, 2007, and ClustalW (1.82) multiple sequence alignment European Bioinformatics Institute 53 were used for sequences alignments of antifungal peptides. T-Coffee is a multiple sequence alignment package 54. Given a set of sequences (Proteins or DNA), T-Coffee generates a multiple sequence alignment. After sequence alignment, some correlations between certain sequences were found, which have been repeated in all of antifungal peptides in each group. AFPs with more sequence homology were selected to extract final pattern among them using CLASTALW alignment. Peptidomimetic structures were designed based on the final pattern obtained from final sequence alignment using T-coffee in which scores are based on colours; positions that have no consistency with the in-house peptide library are in blue, a little in green, better positions in yellow, then orange, and finally red. Yellow to red positions are expected to be entirely correct.
Similarity search was done to find close amino acid sequences to final pattern from sequences alignments in order to find crystallography structures. In the case of our study, T-coffee sequence alignments showed scores between average and good by having dark orange to red colours. The Basic Local Alignment Search Tool (BLAST) 55, and RCSB Protein Data Bank 56 were used to achieve this goal.
Mimetic design: In order to design mimetic counterparts, SuperMimic software 57,58 was used. This tool is being used for finding potential non-peptidic building blocks that can replace or mimic parts of a protein, or peptide and conversely for identifying locations within a protein where such building blocks can be inserted. It identifies compounds that mimic parts of a protein, or positions in proteins that are suitable for inserting mimetics. SuperMimic provides a library of 126 peptidomimetic structures which have been arranged in sublibraries such as beta-turn- or gamma-turn-mimetics 59. SuperMimic program © 2006 was used to replace KTC-Y with peptidomimetics structures which are in SuperMimic library.
Peptide and mimetic preparation: The decapeptide KTC-Y (KTCENLADTY) was synthesized and purchased from SBS biocompany, Beijing, China. Compounds C1 and C2 were purchased from Sigma Aldrich, Eteinhim, Germany, and C3 and C4 purchased from Enamine chemical supplier company.
Protoplast preparation: Protoplasts were prepared according to the protocol described by Osherov and Romano 57 with some modifications. Briefly, 20 ml of Sabouraud dextrose broth medium was inoculated with 109/ml fresh spores of Aspergillus niger (A. niger) N402 or Aspergillus fumigatus (A. fumigatus) Af293 and incubated for 6-7 hr in a shaker incubator, 32oC, 250 rpm until more than 70% of conidia formed germ tube. Germinated spores were then harvested and resuspended in a lytic enzyme mix (5% Glucanex in 0.6 M KCl, 0.05 Citric acid pH=5.8, 1% glucose) and incubated for 2 hr at 30oC, 100 rpm. Resulting protoplasts were harvested by centrifugation for 5 min at 2000 rpm in a bench top centrifuge. Protoplasts were washed twice in 20 ml 0.6 M KCl and resuspended in 1 ml of RPMI 1640 medium, prepared without bicarbonate, buffered with 0.165 M 3-(N-morpholino) propanesulfonic acid (MOPS), and adjusted to pH=7.0 with 10 M NaOH. This medium also contained 2% glucose and 1.2 M sorbitol as an osmotic stabilizer. Antifungal peptide was obtained in powder form and a stock solution of 5 mg/ml in water was prepared. MIC level was determined in 96-well microtiter plates through preparation of twofold serial dilutions across the concentration range (0-200 µg/ml) of antifungal peptide in the above medium. Total volume of each well was 100 µl containing 104 fungal cells. The plate was covered and incubated at 37°C for 24-48 hr. The MIC values were obtained by reading the concentration of the well with no growth.
Antifungal screening: A. niger N402, Candida albicans ATCC 10231 and Saccharomyces cerevisiae PTCC 5052 were used as test strains. The compounds were dissolved in Dimethyl Sulfoxide (DMSO) to reach a concentration of 10 mg/ml. The broth microdilution method was performed for antifungal activity tests. The absorbance was read at 530 nm for fungi inoculums to reach the suitable density of microorganisms, equal to 103 fungi in each well of 96-well microplate after the final dilutions. Working fungal culture was prepared from the stock fungal culture, a 1:1000 dilution with broth (e.g.10 µl stock fungal culture: 10 ml broth). Sabouraud Maltose Broth (SMB) was used as medium.
Modified antimicrobial susceptibility testing based on NCCLS M27-A method was performed 60,61. Broth (100 μl) was added to each well of a 96-well plate and then 40 µl of compounds and 60 µl broth were added to well (A), then a solution (100 μl) serially diluted from well (A) by taking 100 µl into (B) was obtained. This two-fold dilution was continued down the plate and 100 µl from the last well (H) was discarded. Then all the wells were filled with 100 µl of working yeast culture. Itraconazole was used as a reference in fungi test. For this experiment the following controls were prepared: wells containing serial dilution of DMSO and itraconazole only. The plate was covered and incubated at 37°C for 24-48 hr. The MIC values were obtained by reading the concentration of the well with no growth.
Results :
Sequences alignments: Various sets of sequence alignments were carried out in order to find better homology among antifungal peptide sequences (Table 1). The final peptide pattern which was designed in our study and led to KTC-Y was obtained from 10 antifungal peptides that showed more sequence homology to each other with high score -using T-COFFEE- shown in figure 1. The final sequence alignments pattern was K-CENLA-DTY in which "-" could be any amino acid. Source of the all 10 AFPs which were used for the final sequence alignments and the final pattern obtained from them was originally from plants. Hence, we obtained a pattern close to defensin sequence which is AFP from plant kingdom. This pattern was used in similarity search to find crystallography structures close to it. WU-BLAST2 - Protein Database Query, NCBI-BLAST2 - Protein Database Query, FASTA and SSEARCH - Protein Similarity Search, and RCSB Protein Data Bank were used to discover the structures. Interestingly, a structure with similarity of 90.909% to our final pattern was found which its PDB ID is 1JKZ.
Information about this protein and its structure as indicated in Protein Data Bank (PDB) included, molecular description DEFENSE-RELATED PEPTIDE 1; classification: antifungal protein; source scientific name: Pisum sativum; family: plant defensins. The final peptide pattern obtained from multiple alignment led to the structure that its source was from plant, and belonged to defensins family. Having high sequence identity to 1JKZ, starts from N-terminal part of peptide which is important part of peptide for its antifungal activity.
In this study, it was necessary to input just N-terminal part of crystallized structure of 1JKZ (which contains 10 amino acid sequences with high similarity to final pattern) into SuperMimic program and the other parts of this structure were not needed. Therefore, it was needed to separate the mentioned area (N-terminal) from the other parts of its structure. Swiss-pdb Viewer 62 was used to cut N-terminal sequences of our candidate in order to obtain the 3D structure of related segment. This structure named KTC-Y and was used as an input structure in SuperMimic program in order to obtain peptidomimetic structures.
Mimetic design: About 114 peptidomimetics structures were matched to place in KTC-Y using SuperMimic program. Peptidomimetics with lower than 9×10-3 Å Root Mean Square Deviation (RMSD) and more similarity to KTC-Y back bone and its side chains were selected. KTC-Y was divided into 2 parts to study and determine potent antifungal region of its structure between its N-terminal and C-terminal. Four peptidomimetics structures were obtained; two structures were designed for its N-terminal part and two structures for its C-terminal part (Figure 2). Two peptidomimetic structures were generated by replacing of the back bone and side chains of N-termini domain of KTC-Y, named A1 and A2 with RMSD equal to 0.009 and 0.007 Å, respectively. Molecules B1 and B2 represent peptidomimetics structures from C-terminal part of KTC-Y that contains amino acids of 5 to 10 and resulting RMSD equal to 0.008 and 0.007 Å, respectively. These structures are not small enough to possibly pass through the fungal membrane.
Consequently, similarity search was done to find smaller structures with high similarity to the four peptidomimetics. Using Enhanced NCI Database Browser 63, this database also shows summary of data about structures and demonstrates that whether the selected structure has been studied previously or not. This database can also predict activity of structures and can help to estimate the activity of structure against microorganisms. Among all structures that were found, structures that had not been studied before for antifungal activity were selected. Zinc 64 as a database of commercially-available compounds for virtual screening was applied to uncover these compounds availability through companies for purchase. Finally, four structures were chosen among the resulting molecules for antifungal bioactivity screening (Figure 3).
Protoplast and antifungal screening: Four compounds that were obtained through similarity search (Figure 3) were screened for their protoplast and antifungal properties. These compounds were tested to identify the level of inhibition and the results are shown in table 2. The general ability of the test was affirmed by including a positive control agent. There was no inhibition by DMSO solvent as a negative control. Among the compounds tested, C3 and C4 did not show acceptable antifungal activities. C1 and C2 had more antifungal activities against A. niger and Saccharomyces cerevisiae (S. cerevisiae). Compound C2 represents the least MIC among the compounds against A. niger and S. cerevisiae. Compound C2 is more effective against A. niger than Candida albicans (C. albicans) and shows more antifungal activity on the tested fungal strains than C1, C3 and C4. Due to these results C2 and C1 could be an option for future studies and developmental research.
The decapeptide KTC-Y was tested in antifungal bioassay and showed no activity up to 2000 µg/ml in neither of fungal strains assayed; it means that higher concentrations of KTC-Y should be used in order to detect its fungal inhibitory effect or the peptide cannot pass through the cell membrane. To test the second possibility, when KTC-Y was tested against A. fumigatus in the protoplast assay, it showed a potent activity, MIC=25 µg/ml (Table 2). The above-mentioned test provides further support for the possibility that KTC-Y peptide is active and it might be the cell wall of A. fumigatus that does not allow KTC-Y penetration into its active site on the cell membrane or in the cell itself.
Discussion :
An alarming rise in life-threatening systemic fungal infections of varying types and frequencies is seen in immunocompromised patients such as AIDS, cancer and transplant patients. Invasive medical procedures such as the use of vascular catheters, peritoneal dialysis, haemodialysis and parenteral nutrition have also contributed to the rise in fungal infections. The problem is made worse by the emergence of fungal strains that are resistant to currently used antifungal agents. Therefore, the design of new drugs and further improvement of currently used drugs are essential. The searches for appropriate cellular targets as well as the complete biophysical characterisation of these molecules have begun.
Recent, advances in cell and molecular biology, microbiology, biochemistry and recombinant DNA technology have greatly contributed to the characterisation of molecular targets. The modern approach to drug discovery focuses on identifying and describing biochemical targets – mainly proteins – and subsequently matching those targets with small molecules that have the properties needed to inactivate them, i.e. structure-based drug design.
Three-dimensional characterisation of antifungal drug targets and understanding the mode of action of antifungal agents are indispensable tools in modern drug research. The success of structure-based drug design demands an iterative procedure 65. The structure of the protein analyzed in detail allows the design of a ligand that must then be synthesised and tested. Biological data and new protein-ligand complex determination help to refine working hypotheses about complementarity. Repeating these steps leads to incremental improvement in the initial compound.
Proteins achieve their functions by folding into compact, well-ordered structures 66. Multiple alignments of protein sequences are important in many applications, including phylogenetic tree estimation, secondary structure prediction and critical residue identification 67. Sequence alignment is a common tool in bioinformatics and comparative genomics 68. In the current study, investigation for extracting pattern with homology among all AFPs amino acids sequences using ClustalW, BLAST, and T-Coffee led to final pattern with 10 amino acids sequences. AFPs that showed more homology to this pattern belonged to plants. Crystallized structure that belonged to defensins group was obtained from this pattern. Defensins are major AFPs from plant 69. Combining these results and information together, and with the help of sequence alignments, we could realize that final pattern are from antifungal peptides originating in plants, and therefore, the crystallized structure extracted similar to KTC-Y structure, belongs to defensins that are most important AFPs in plants. Pleurocidin is a 25-residue peptide which has been studied for its interaction with membrane of human pathogenic fungus C. albicans. It was shown that this peptide exerted fungicidal activity against C. albicans with the disruption of plasma membrane 70. Piscidin is another example of cationic peptide which has been investigated for its antifungal activity, being able to permeabilize the model phospholipids membranes 71.
Design of mimetic molecules has been reported previously. For example, Fullbeck et al 59 used SuperMimic software in order to be able to present folding of the 35-residue subdomain (H35) at its C-terminus of the villin headpiece by irradiation, to replace parts of its main chain without changing the overall structure of the subdomain. Our efforts to design peptidomimetics structures led to finding four compounds through in silico analysis. Antifungal assay results indicated that C2 posses more antifungal activity against selected microorganisms C. albicans and S. cerevisiae, and A. niger rather than the others compounds. The question that we faced was that why this compound is more potent for its antifungal activity or whether this compound acts on lipid bilayer membrane of tested microorganisms.
Looking into this compound’s structure, it is realized that this structure consists of tryptophan which could affect its antifungal activity. Therefore, a literature survey was carried out to discover influence of tryptophan on antifungal activity. Tryptophan is generally not an abundant amino acid residue in peptides or proteins. The effect of tryptophan derivative here might be corresponding to its behaviour in AMP in which the partitioning of peptides into membranes is imposed by its propensity to position itself near the membrane/water interface. Examples of antimicrobial peptides exist that are rich in tryptophan include tritrpticin (VRRFPWWWPFLRR) and indolicidin (ILPWKWPWWPWRR-amide) 72. Indolicidin exhibits potent antifungal and antiviral activities that is rich in tryptophan residues 73. Lawyer and his colleagues 74, found that a 13 amino acid tryptophan-rich region with the sequence of VRRFPWWWPFLRR had strong antimicrobial activity with a wide spectrum.
Tryptophan residues play a crucial role in membrane spanning proteins as well, has a strong preference for the interfacial regions of lipid bilayers. In addition, tryptophan residues are involved in protein folding, forming both native and non-native hydrophobic contacts even in denatured proteins to ensure their proper folding. Further testament to the significance of tryptophan is the fact that upon screening of a complete combinatorial library of hexapeptides, peptides rich in these two amino acids (W and R) were found to possess the highest antimicrobial activities 75.
Ramamoorthy et al 76, investigated the correlation between cell selectivity and membrane interactions of G15, which is antimicrobial peptide granulysin, and discovered the role of interaction of tryptophan residues and possibly the hydrophobic sidechains with the phosphate head groups for a tight binding of the G15 to the lipid bilayers. They suggested that the tryptophan residue is located at the bilayer interface and not deep inside the hydrophobic region of the membrane. Andreu and colleagues 31, during study on cecropins suggested that AMPs containing more tryptophan are being active against microorganisms.
The mimetic design has many applications in biochemical studies for development of new therapies and also pest control 78,79. Recently, some halide substituted indole dihydropyrimidines were synthesized and their antifungal activities have been reported 80. Singh and colleague 81 have isolated an indole alkaloid Alstonia venenata named venenatine, which shows antifungal activity against some plant pathogenic and saprophytic fungi.
Conclusion :
In this study, we investigated a group of antifungal peptides through bioinformatics investigation that could extract a general pattern to be in common in the N-terminal region. After further analysis, the KTC-Y peptide was chosen to be synthesized. In our preliminary studies, the mentioned decapeptide showed to be active only in protoplast cells, which is an indicator of deep target site for such molecule and role of cell wall as a penetration barrier for this peptide. By computational tools and design of mimetics of and generation similar to mimetics, we were able to reach a group of molecules that are active and can be used for further studies in antifungal drug discovery and design processes. Further experiments on such compounds to determine their mode of action and synthesis of their derivatives are currently in place in our group.
Acknowledgement :
The authors would like to thank Mrs. S. Enayati for her useful technical assistance.

Figure 1. Final sequences alignment among AFPs that led to the selected pattern
|
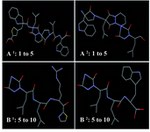
Figure 2. Peptidomimetic structures that designed by SuperMimic Program
|
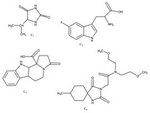
Figure 3. Final compounds for antifungal screening, named C1, C2, C3 and C4
|

Table 1. Antifungal peptides, their amino acids sequences, molecular weight and their sources
Table 1. Antifungal peptides, their amino acids sequences, molecular weight and their sources (Continued)
|
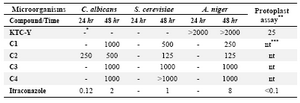
Table 2. Protoplast assay result of KTC-Y against A. fumigatus Af293, and antifungal effect of four mimetic compounds against A. niger, C. albicans, and S. cerevisiae, as obtained by the broth dilution method; MIC values are expressed in µg/ml
*"-" means no readable growth; ** The test was performed on A. fumigatus; *** "nt" means not tested
|
|