Construction and In vitro Expression Analyses of a DNA Plasmid Encoding Dense Granule GRA5 Antigen of Toxoplasma gondii
-
Babaie, Jalal
-
Molecular Parasitology Laboratory, Pasteur Institute of Iran, Tehran, Iran
-
 Golkar, Majid
Ph.D., Pasteur Institute of Iran, Tehran, Iran, Tel: + 98 21 66968855 Fax: + 98 21 66492595 E-mail: golkar@pasteur.ac.ir
Golkar, Majid
Ph.D., Pasteur Institute of Iran, Tehran, Iran, Tel: + 98 21 66968855 Fax: + 98 21 66492595 E-mail: golkar@pasteur.ac.ir
-
Molecular Parasitology Laboratory, Pasteur Institute of Iran, Tehran, Iran
Abstract: Toxoplasmosis is an infection caused by the protozoan parasite Toxoplasma gondii (T.gondii) throughout the world. Although usually asymptomatic, the infection can cause serious medical problems in immunocompromised individuals and fetuses. Toxoplasmosis also causes considerable economic loss because of abortion in livestock. DNA vaccination is a promising approach against intracellular parasites such as T.gondii. The goal of this study was to construct and evaluate functionality of a mammalian plasmid expressing GRA5 anti-gen of T.gondii as a possible DNA vaccine. GRA5 gene fragment, devoid of the signal sequence, was amplified from genomic DNA of T.gondii RH strain, and cloned into pcDNA3.1 plasmid. The pcDNA3.1-GRA5 (pGRA5) was analyzed by restriction enzyme digestion followed by sequence determination. The pGRA5 was transfected into HEK 239-T human kidney cells, and expression of GRA5 antigen was investigated by Western blotting and immunofluorescence staining. The sequence encoding GRA5 was cloned into pcDNA3.1 plasmid. Restriction digestion of pGRA5 with Pst I enzyme showed correct insertion of GRA5 DNA into the plasmid. Sequence analysis revealed 100% homology with the published sequence of gra5. Immunofluorescence and Western blotting analyses of HEK 293-T cells transfected with pGRA5 showed specific expression of GRA5. Immunogenicity of pGRA5 will be evaluated in mice.
Introduction :
Toxoplasma gondii is a common, single-celled parasitic agent which causes Toxoplasmosis worldwide. It is estimated that up to one-third of the world human population are infected by the parasite (1). The infection is transmitted orally by ingesting parasite cysts in undercooked meat, or by accidental ingestion of oocysts shed in the feces of the feline (2-4). Toxoplasma infection is often benign in immunocompetent individuals, but in individuals with impaired immune system, such as AIDS patients, toxoplasmosis is an important opportunistic infection. In these people, latent T.gondii infection can be reactivated to cause fatal Toxoplasmic encephalitis (5). Primary infection during pregnancy can result in permanent neurological and ocular complications in the fetus (3). In livestock, particularly in sheep and goats, infection can lead to abortion, causing significant loss to the farming industry.
Chemotherapy efficiently controls acute Toxoplasma infection and reduces rate of neurological and ocular problems in the congenitally infected newborns, however, it cannot prevent development of latent infection as represented by tissue cyst production in neural and muscular tissues (3-6). Therefore, it would be of great interest to produce an effective Toxoplasma vaccine to prevent the infection. So far, the only developed vaccine is the live, attenuated tachyzoite S48 strain known as Toxovax (7), however, it is not widely accepted because of its adverse effects, short shelf-life, and high cost (8-9). Live vaccines also carry a risk of accidental infection of humans and unexpected harmful reverse mutations. Subunit vaccines, derived from native antigens of the parasite as recombinant protein or DNA, might overcome these hurdles (9,10).
Vaccinations by DNA-based structures have been shown to elicit a potent, long-lasting humoral and cell-mediated immunity, in addition to providing protection against viral, bacterial, and parasitic infections (11-14). The ability of DNA vaccination to produce the encoded protein inside the cell makes it an attractive immunization approach against T. gondii (15-18). Numerous studies showed immunogenicity and protective efficacy of dense granule (GRA) antigens (9). GRA antigens are stored within Toxoplasma dense granules and secreted into the Parasitophorous Vacuole (PV) after parasite invasion (19,20). DNA vaccination with GRA antigens induced strong immune responses and protected against Toxoplasma infection in different animal models (9,18,21-24).
GRA5 is a small protein of 120 amino acids containing a signal peptide of 25 aminoacids, two hydrophobic regions, and a single α-helical transmembrane domain (25). It was found in both tachyzoites and bradyzoites life stages of T.gondii (26). Holec et al reported serodiagnostic utility of GRA5 for detection of T.gondii IgG antibodies (27). Immunization with native GRA5 (28) or an antigenic cocktail containing recombinant GRA5 (29) induced protective immunity against T.gondii infection. In this study, a recombinant plasmid encoding mature full-length GRA5 antigen was constructed and tested for expression in a mammalian cell line, as a basis toward developing a DNA vaccine against Toxoplasmosis.
Materials and Methods :
Parasites
Tachyzoites of virulent RH strain of T. gondii were injected into peritoneal cavity of Swiss mice. Three days later, tachyzoites were harvested, washed with phosphate-buffered saline (PBS) and stored at -80 oC until used.
Reagents
Escherichia coli DH5α (Invitrogen, Carlsbad, CA) strain bacteria were used for cloning purpose. The mammalian expression plasmid pcDNA3.1/Hygro (24) (Invitrogen, Carlsbad, CA) was used for expression of GRA5.
PCR amplification of GRA5 gene
The GRA5 gene fragment encoding mature full-length protein, residue 27-120, was PCR- amplified from genomic DNA of T. gondii RH strain using the following oligonucleotide primers: Forward primer: 5 ́ GCC ACC ATG GGT TCA ACG CGT GAC GTA GGG TCA G 3́
Reverse primer: 5 ́ GCT GAG ACA ACA AAG AGC CGA GCA ACA CAG TGC 3́
The complete Kozak translational consensus sequence (underlined) was introduced in the beginning of the PCR product. PCR amplification was performed using the following conditions: 1 cycle of 95 °C for 5 min then 30 cycles of 95 °C for 30 sec, 58 °C for 1 min, and 72 °C for 30 sec. Final primer extension was extended to 30 min at 72 °C. PCR product was analysed by electrophoresis on 1.0 % agarose gel.
Cloning of gra5
The PCR product was inserted into T/A cloning vector pTZ57R/T, excised by KpnI / ApaI enzymes, and subcloned into KpnI/ApaI restriction sites of pcDNA3.1/Hygro (24) plasmid. Selection of recombinant clone harboring GRA5 DNA was performed using PstI digestion of recombinant plasmids followed by sequence determination. The resulting plasmid was named pGRA5.
Transient transfection of HEK 293-T cells
HEK 293-T cells, an embryonic kidney cell line, were grown at 37 °C in a humidified 5% CO2 atmosphere in 35-mm wells in Dulbecco’s Modified Eagle Medium (DMEM) containing 100 U/mL each penicillin and streptomycin and 10% fetal calf serum, and were transfected at 50-70% confluency with 4 µg of pGRA5 or pcDNA3.1 using PolyFect transfection reagent (Qiagen, Hilden, Germany) according to the manufacturer instructions. The transfected cells were incubated with PolyFect for 16 to 24 hr before replacing the medium with 2 ml of fresh medium. After 3 days, cell monolayers were washed 3 times with Phosphate Buffered Saline (PBS) and scraped into 1 ml of PBS. Cells were then recovered by centrifugation at 12,000 × g for 15 min and stored at -20 °C.
Immunofluorescence staining
HEK 293-T cells were grown on 12-mm coverslips in 24-well plates and transfected with pGRA5 as described in the previous section. Three days after transfection, cells were fixed with 5% paraformaldehyde in PBS for 10 min, and permeabilized with 0.2% Triton X-100 in PBS for 3 min. Subsequently, cells were incubated with mAb anti-GRA5 TG17-113 (30), diluted 1:500 in 1% FBS in PBS for 1 hr at room temperature. Cells were washed with PBS and incubated for 1 hr with FITC-conjugated anti-mouse IgG (Sigma, St. Louis, MO, USA), diluted 1:20,000 in 1% FBS in PBS. Coverslips were mounted using Prolong antifade reagent (Molecular Probes) and observed using a Zeiss Axioplan II equipped for phase-contrast and epifluorescence microscopy. Photographs were taken at the magnification × 100 using a Zeiss Axiocam MRm coupled to the AXIOVISION 4.5 software and processed with ADOBE PHOTOSHOP 6.0.
Western blot analysis
Western blot analysis of HEK 293-T cells
transfected with pGRA5 was performed on cell pellets of single 35-mm wells. The pellet was resuspended in 50 µL of SDS-PAGE sample buffer, sonicated, boiled for 5 min, and 15 μl was loaded onto a 15% polyacrylamide gel. Proteins were transferred onto nitrocellulose membrane via electrophoresis, carried at 100 V and 350 mA for 1 hr, using Bio-Rad transfer system (Bio-Rad, Hercules, CA). The membrane was saturated for 1 hr with 5% fat-free dried milk in PBS and probed with the mAb anti-GRA5 diluted 1:1000 in saturation buffer. The membrane was incubated for 1 hr with peroxidase-conjugated goat anti-mouse IgG (Sigma, St. Louis, MO, USA) diluted 1:20,000 in saturation buffer, and signals were detected using super signal ECL (Enhanced Chemiluminescence) system (Pierce chemical, Rockford, IL).
Results :
Cloning of gra5
The DNA encoding mature GRA5 antigen, residue 27-120 was amplified from genome of T.gondii RH strain by means of PCR. The complete Kozak consensus sequence, GCC ACC ATG G, was introduced in 5 ́ of the GRA5 DNA. The specific product of 333 bp was observed in gel agarose electrophoresis (Figure 1) The PCR product was inserted into T/A cloning vector, pTZ57R/T, excised by KpnI/ApaI enzymes and subcloned into KpnI/ApaI restriction sites of pcDNA3.1 mammalian expression plasmid. Screening of recombinant clones harboring pcDNA3.1-GRA5 (pGRA5) plasmid was performed by restriction digestion using PstI enzyme. Digestion of the recombinant plasmid produced two fragments of 4243 and 1626 bp, but nonrecombinant plasmid gave rise to only one DNA band (Figure 2). Sequence analysis of the cloned gene revealed 100% homology with the published sequence of gra5.
Expression of GRA5 protein in mammalian cells
Expression of the recombinant protein by mammalian cells transfected with a plasmid DNA is required for stimulation of the immune system. As such expression of GRA5 protein was assessed in HEK 293-T cells transfected with pGRA5. Transfected cells were cultured for 3 days, washed, and expression of recombinant GRA5 was assessed by Western blotting and immunofluorescence staining. The result showed that HEK 293-T cells transfected with pGRA5 produced antigenic GRA5 protein (Figure 3). In Western blotting, recombinant GRA5 was detected as a band of about 18 kDa, which was about the same size as the native protein (Figure 3). In contrast, no immunoreactive material was found in the lysate of HEK 293-T cells transfected with pcDNA3.1 plasmid. The smaller and larger protein bands in the lane 4 are probably T.gondii antigens reacted non-specifically with the mAb anti-GRA5. Immunofluorescence staining showed HEK 293-T cells transfected with pGRA5 specifically produced GRA5 protein (Figure 4.) which was located in the cytoplasm.
Discussion :
The DNA encoding GRA5 protein was amplified from RH strain of T.gondii and cloned in the pcDNA3.1 mammalian expression plasmid. Expression of GRA5 antigen in HEK 293-T cells transfected with pGRA5 was investigated to make sure if the recombinant plasmid is able to direct GRA5 production upon DNA vaccination of mice.
The first step in making a DNA vaccine is to make a recombinant plasmid encoding the desired antigen followed by investigating functionality of the plasmid construct. We cloned GRA5 DNA in pcDNA3.1 plasmid under transcriptional control of a strong cytomegalovirus (31) early promoter. The introduction of complete Kozak sequence in the beginning of GRA5 gene would allow optimum initiation of protein expression (32). Transfection of human embryonic kidney cells, HEK 293-T cells, with pGRA5 plasmid resulted in specific production of GRA5, as shown by Western blotting and immunofluoresnce staning.
Gendrin et al (33) transiently expressed full-length GRA5 (including the signal peptide) in HEK 293-T cells and addressed the localization of GRA5. They found that GRA5 behaves like an integral membrane protein with a type I topology in mammalian cells. They also showed that GRA5 N-terminal is necessary and sufficient to mediate membrane insertion into the PV membrane (33). In this study, we observed that GRA5 without the putative signal peptide was localized to the cytoplasm of HEK293-T cells. It was expected as the presence of signal peptide is necessary for entry in to the endoplasmic reticulum and subsequent membrane insertion.
We aimed at making a GRA5 DNA vaccine as previous studies suggested immunogenicity and vaccine potential of GRA5 antigen. The rate of transplacental infection in rats injected with Incomplete Fround’s Adjuvant (IFA) was 41.7%, however, it was reduced to 15% in rats immunized with FPLC-purified GRA5 formulated in IFA (28). Igarashi et al (29) showed both local and systemic immune responses in BALB/c mice immunized with a mixture of recombinant proteins GRA5, GRA7 and ROP2 combined with cholera toxin. This mixture offered about 60% protection against brain cyst formation (29).
Conclusion :
In conclusion, this study reported construction of a plasmid DNA encoding GRA5 protein of T.gondii. We confirmed that pGRA5 plasmid is able to direct synthesis of antigenic GRA5 protein in mammalian cells. The ability of pGRA5 for induction of a protective immune response will be evaluated in mouse models of toxoplasmosis.
Acknowledgement :
We would like to appreciate Drs. Marie-France Cesbron-Delauw and Corinne Mercier for providing some reagents and reviewing the manuscript. This work was supported by the research grant No. 256 from Pasteur Institute of Iran.
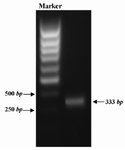
Figure 1. PCR amplification of GRA5. A pair of specific primer was designed and used to amplify GRA5 gene from genome of T.gondii RH strain. PCR product was analyzed by 1% gel agarose electrophoresis
|
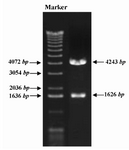
Figure 2. Screening of recombinant clones containing GRA5 gene. GRA5 PCR product was inserted into pcDNA3.1 plasmid and the recombinant plasmid, pGRA5, was transformed into DH5α bacteria. Screening of recombinant clones was performed by PstI restriction digestion. Digestion of recombinant plasmid harboring GRA5 gene resulted in two DNA fragments of 4243 and 1626 bp
|
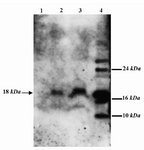
Figure 3. Western blot analysis of expression of GRA5 in HEK 293-T cell. Expression of the GRA5 protein was evaluated in HEK 293-T cells transfected with pGRA5. The recombinant protein was probed with the mAb anti-GRA5 TG17-113 and GRA5 was found as a band of about 18 kDa, which was about the same size of the native protein (Lanes 2 and 3). Untransfected HEK 293-T cells were used as the negative control (Lane 1) and takyzoites of RH strain were used as the positive control (Lane 4)
|
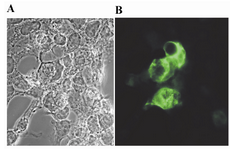
Figure 4. Immunofluorescence expression analysis of GRA5 in HEK 293-T cells transfected with pcDNA3.1 (A) or pGRA5 (B). At 72 hr post-transfection, the cells were fixed and immunofluorescence staining was performed using a mAb anti-GRA5 followed by FITC-conjugated anti-mouse IgG. Fluorescent images were examined
|
|