Clinically Significant Dysregulation of hsa-miR-30d-5p and hsa-let-7b Expression in Patients with Surgically Resected Non-Small Cell Lung Cancer
-
Hosseini, Sayed Mostafa
-
Department of Genetics, Faculty of Biological Sciences, Tarbiat Modares University, Tehran, Iran
-
 Mohammad Soltani, Bahram
Department of Molecular Genetics, Faculty of Biological Sciences, Tarbiat Modares University, Tehran, Iran, Tel: +98 21 82883464, E-mail: soltanib@modares.ac.ir
Mohammad Soltani, Bahram
Department of Molecular Genetics, Faculty of Biological Sciences, Tarbiat Modares University, Tehran, Iran, Tel: +98 21 82883464, E-mail: soltanib@modares.ac.ir
-
Department of Genetics, Faculty of Biological Sciences, Tarbiat Modares University, Tehran, Iran
-
Tavallaei, Mahmoud
-
Human Genetic Research Center, Baqiyatallah University of Medical Sciences, Tehran, Iran
-
Mowla, Seyed Javad
-
Department of Genetics, Faculty of Biological Sciences, Tarbiat Modares University, Tehran, Iran
-
Tafsiri, Elham
-
Department of Molecular Medicine, Biotechnology Research Center, Pasteur Institute of Iran, Tehran, Iran
-
Bagheri, Abouzar
-
Department of Clinical Biochemistry and Genetics, Molecular and Cell Biology Research Center, Faculty of Medicine, Mazandaran University of Medical Sciences, Sari, Iran
Abstract: Background: The cyclin E2 (CYCE2) is an important regulator in the progression and development of NSCLC, and its ectopic expression promoted the proliferation, invasion, and migration in several tumors, including Non-Small Cell Lung Cancer (NSCLC). However, the upregulation of CYCE2 in NSCLC cells suggested that it has a key role in tumorigenicity. In addition, the RAS family proteins as oncoproteins were activated in many major tumor types and its suitability as the therapeutic target in NSCLC was proposed. Considering the crucial role of microRNAs, it was hypothesized that altered expression of hsa-miR-30d-5p and hsa-let-7b might provide a reliable diagnostic tumor marker for diagnosis of NSCLC.
Method: Real-time RT-PCR approach could evaluate the expression alteration of hsa-miR-30d-5p and hsa-let-7b and it was related to the surgically resected tissue of 24 lung cancer patients and 10 non-cancerous patients. The miRNAs expression was associated with clinicopathological features of the patients.
Results: Hsa-miR-30d showed a significant downregulation (p=0.0382) in resected tissue of NSCLC patients compared with control group. Its expression level could differentiate different stages of malignancies from each other. The ROC curve analysis gave it an AUC=0.73 (p=0.037) which was a good score as a reliable biomarker. In contrast, hsa-let-7b was significantly overexpressed in tumor samples (p=0.03). Interestingly, our findings revealed a significant association of hsa-let-7b in adenocarcinoma tumors, compared to Squamous Cell Carcinomas (SCC) (p<0.05). Also, analysis of ROC curve of hsa-let-7b (AUC=0.74, p-value=0.042) suggests that it could be as a suitable biomarker for NSCLC.
Conclusion: Together, these results suggest a possible tumor suppressor role for hsa-miR-30d in lung tumor progression and initiation. Moreover, upregulation of hsa-let-7b was associated with the tumor type.
Introduction :
Lung cancer is the most common reason of cancer-related mortality for patients’ suffer from cancer in Iran. Owing to life style change, air pollution and increasing tobacco consumption, incidence of lung cancer is growing and it is one of the five common cancer types in Iran 1.
Non-Small-Cell Lung Cancer (NSCLC) is responsible for about 80-85% of all cases of lung cancer 2. Late diagnosis of lung cancer is one of the key causes for high mortality 3. Moreover, the prognosis of patients with end-stage of NSCLC remains unfavorable. By means of presently existing methods, more than around ~75% of lung tumor are diagnosed at untreatable stages 2,4,5.
MicroRNAs (miRNA) are endogenous short class of small non-coding RNAs (18-24 nucleotides) which fine-tuning gene expression dosage through targeting mRNAs 6. MiRNAs regulate an extensive array of biological processes, such as cell proliferation, adhesion, apoptosis, cell death and differentiation 7.
The let-7 family, as founding member of miRNAs, is considered as tumor suppressors via negatively regulating RAS oncogenes in NSCLC 8, with downregulated expression correlating with poor prognosis. Aberrant expression of the family members can however promote the reprogramming of these normal processes and motivate cell proliferation and tumor metastasis 9,10. The let-7 family was previously evaluated as lung reprogramming-related miRNAs, with aberrant expression levels which promoted NSCLC carcinogenesis.
Several reports indicated that hsa-hsa-miR-30d is negatively regulated in both NSCLCs 11. hsa- miR-30d is located at 134804876 bp to 134804945 bp on chromosome 8q24.22. It belongs to the has-miR-30 family including hsa-miR-30a-f that keeps a sequence highly conserved between species.
Yao et al found that hsa-miR-30d was upregulated and involved in metastasis process via targeting GNAI2 in hepatocellular carcinoma 12. Extensive research has shown that hsa-miR-30d was deregulated and functions as a tumor suppressor in many tumors, including prostate cancer 13-15, hepatocellular carcinoma 12 and medulloblastoma 16.
Accordingly, miRNA alteration investigation could be used to classify tumors based on their stage of malignancies. miRNAs profile analysis, only or in combination with traditional methods, potentially have the ability to increase chance of diagnosis, prognosis, identifying effectiveness predictive biomarkers and finding more effective molecular targets for novel therapeutic procedures 5,15. However, the clinical pathologies feature the role of hsa-let-7b and hsa-miR-30d in the tumorgenesis of NSCLC.
Considering the ectopic expression of cyclin E2 (CYCE2, Gene ID: 9134) in lung tumor tissues and its role in cell cycle G1/S transition and proliferation, it was hypothesized in this study that the expression of hsa-miR-30d-5p which targets cyclin E2 might be potentially applied as a tumor marker in NSCLC. In addition, one of the signal transduction pathways related to NSCLC are members of the RAS GTPase family, which contain multiple putative let-7 binding sites. Using bioinformatics approaches, a putative RAS-targeting microRNA (hsa-let-7b) was predicted, and then the expression levels of hsa-miR-30d and hsa-let-7b in NSCLC tumor samples versus non-tumor tissues were evaluated. Moreover, an attempt was made to investigate whether expression levels in their expression alteration were associated with the clinicopathological features of tumor including staging, smoking status, and type of lung tumor.
Materials and Methods :
Study design and clinical samples: In this study, the sample included 24 patients with surgically resected NSCLC and 10 matched distant noncancerous tissues from Baqiyatallah Hospital, Tehran, obtained between January 2014 and May 2016. All the patients included in our study had to meet the following criteria: (i) patients must be associated with the diagnosis of NSCLC and (ii) gold standard techniques, comprising lung biopsy samples and imaging procedures, were applied to confirm the histopathological features and tumor stages of NSCLC patients. The exclusion criteria were: (i) none of the patients has ever received radiotherapy or chemotherapy; (ii) there are no significant differences in age, gender and smoking status; and (iii) patients without sufficient data. Ten healthy controls were selected including smokers and non-smokers but with no history of pulmonary diseases as a control group, including 2 Benign Pulmonary Nodule (BPN) patients and 8 Chronic Obstructive Pulmonary Disease (COPD).
All patients had undergone flexible bronchoscopy, improved by rapid on-site evaluation performed by an experienced pathologist, at the bronchoscopy unit. The age of the patients was 37-80 years old (mean, 59.29 years) who were 52 male and 16 female. For tissue sample collection, upon removal of the surgical specimens, the tissues were immediately transported to the Pathology Laboratory and the samples were placed in a cryovial, snap-frozen liquid nitrogen for 30 min, and stored at -80 oC until use.
All samples were evaluated immunohistochemically for the expression of two major markers that have been promoted for the classification of NSCLCs. Expression of thyroid transcription factor-1 (TTF-1) was assessed in alveolar type cells and in bronchiolar cells, and its expression is often retained in adenocarcinomas derived from these cell types. Furthermore, p63 immunostaining was performed as a means of discerning squamous differentiation in NSCLC in Baqiyatallah Pathology Laboratory. Written informed consent was achieved and the Ethical and Scientific Committees of Baqiyatallah University of Medical Sciences approved this study. After surgery elimination, all resected tissue specimens were immediately immersed in RNAlater liquid buffer and kept at -80oC until use 17. Smoking history, data of tumor histology, and staging were obtained from patients. The NSCLC patients included 14 adenocarcinomas and 10 SCC, and 9 stage I, 9 stage II, 6 stage III as determined according to WHO classification and the International Union against Cancer staging system. The majority of patients (15/24, 62.5%) were smokers. All the necessary information of patients and healthy controls is provided in table 1.
RNA extraction: Total RNA from the lung tissue specimens was extracted via TRIzol reagent (Ambion) as formerly described 18. The pellet of extracted RNA was dissolved in 15 μl RNA storage buffer (Invitrogen) and kept at -80 oC until use. The RNA yield and A 260/280 ratio were calculated by a NanoDrop spectrophotometer (Thermo Fisher Scientific, Waltham, USA), then 25 fmol of an exogenous synthetic miRNA, UniSP6 (Exiqon, Denmark) as an external control for normalization of sample to-sample differences in RNA isolation process was spiked into each tissue specimen 19.
RNA polyadenylation, cDNA synthesis and quantitation of miRNA expression by RT-qPCR: Before complimentary DNA synthesis, the samples were treated with RNase-free DNase to remove possible traces of genomic DNA. cDNA (Complimentary DNA) synthesis was then performed on 1 µg total RNA based on the poly A tailing method. Initially, a poly A tail was added to the extracted RNAs by incubating 1 μg of RNA 2 μl of 10 x buffer, 0.5 μl of poly A polymerase enzyme, 2 μl of ATP, 0.5 μl of RNase inhibitor (Thermo, Fisher Scientific, Waltham, USA), and DEPC-treated water at 37°C for 10 min. Then, for cDNA synthesis, 2 μl of polyadenylated RNA was mixed with 2 μl of 5 x buffer, 0.5 μl of reverse transcriptase (RT) enzyme, 0.5 μl of specific primers, and 0.5 μl of RNase inhibitor (Ribolock). The mixture was incubated in a thermal cycler at 42°C for 60 min followed by incubation at 85°C for 1 min, to heat-inactivate the reverse transcriptase enzyme.
Real-time quantitative PCR (RT-qPCR) was performed using 1 μl cDNA product, 0.5 μl of specific forward primer, 0.5 μl of universal reverse primer (ParsGenome, Iran), 10 μl of HOT FIREPol EvaGreen qPCR Mix (Solis BioDyne, Estonia) and 8 μl of nuclease-free water (CinnaClon, Iran). The has-mir-16-5p and U6 were used as an internal control. Real time qPCR was performed to measure expressions of miRNAs by using a Step-one plus system (Applied Biosystems, USA) with the following conditions: 95°C for 15 min, followed by 40 cycles of 95°C for 15 s, 61°C for 25 s, and 72°C for 20 s. All RT-qPCR amplifications were performed in duplicate, and a negative control was included for each reaction for quality control.
Statistical analysis: In order to do statistical analysis, firstly, Kolmogorov-Smirnov normality test (KS test) was done to survey the normal distribution of the samples. RT-qPCR data analyses were performed with REST 2009 software (Qiagen, Hilden, Germany) and GraphPad Prism 6 software (GraphPad Software Inc., SanDiego, USA). The results are showed as the mean±standard error of the mean (SEM) from at least three independent trials. Alterations between two groups were analyzed using parametric equations to determine the significance of the observed differences between different groups. All p-values shown were one sided, and a p-value of <0.05 was considered statistically significant. The Receiver Operating Characteristic (ROC) was employed to interpret the optimal sensitivity and specificity levels at which to discriminate normal individuals from NSCLC patients from non-tumor control, and corresponding thresholds were calculated for each miRNA.
Results :
Real-time quantitative PCR analysis and direct DNA sequencing: All sample types were NSCLC tumor type, from which 15 were high grade (II/III), and 9 were low grade. Total RNA was extracted and RT-qPCR was performed via miR-specific primers and outer primer (Table 2) which amplified ~70 base pair products on all samples and the validity of the PCR products were confirmed by agarose gel electrophoresis and direct sequencing.
All data were normalized to the average expression of has-mir-16 and U6, as internal microRNA control. There were no non-specific products or primer-dimer peaks in melt-curve analysis (data not shown).
Hsa-miR-30d is significantly downregulated in NSCLC tissues: Our data showed that there was a significant downregulation of hsa-miR-30d expression levels in NSCLC samples compared to those of their nontumor controls (p<0.0382) (Figure 1A). The patients were separated in two groups including the ones in stages I/II and stage III, and the expression level of hsa-miR-30d significantly decreased in stage III tumors in comparison to that of tumor samples with stage I/II (p<0.0420) (Figure 1B). The levels of hsa-miR-30d expression in NSCLC-smoker group were higher than those in NSCLC-nonsmoker (Figure 1C). Nevertheless, this alteration was also not statistically significant. Similarity, our data showed no statistically significant association between expression levels of hsa-miR-30d in adenocarcinoma in comparison to SCC, which may have been due to the small sample size (24 samples of NSCLC and 10 samples of non-tumors) (Figure 1D).
Hsa-let-7b is significantly upregulated in NSCLC tissues: A significant upregulation of hsa-let-7b expression alterations was observed in tumor samples compared to those of unpaired nontumor controls (p=0.0382) (Figure 2A). The expression alterations of hsa-let-7b were not statistically associated with stages (Figure 2B) and smoking status (Figure 2C). Interestingly, there was a significant upregulation of hsa-let-7b in lung adenocarcinomas, compared to the SCC (p=0.02, Figure 2D).
The specificity and sensitivity of hsa-miR-30d and hsa-let-7b expression levels in discrimination tumor vs. non-tumor samples: ROC curve analyses were conducted to estimate the sensitivity and specificity of the hsa-miR-30d and hsa-let-7b expression levels could discriminate tumor/non-tumor states. As it is evident in figure 3, total area under the curves (AUCs) of hsa-miR-30d and hsa-let-7b were 73 % (CI=0.5264 to 0.9344, p=0.037) and 74% (CI=0.5724 to 0.9167, p=0.042) , respectively.
Discussion :
Recently, remarkable improvements have been developed in the discovery of new biomarkers associated with variations at the molecular level of lung cancer. The researches have demonstrated significant clinical relation in different areas such as diagnosis, response to drugs and tumor classification. Many studies have been concentrated on evaluation expression of key microRNAs involved in NSCLC development, progression and manners of tumor in different stages 20,21.
To investigate the expression alteration of hsa-miR-30d in NSCLC patients, hsa-miR-30d expression was evaluated in various NSCLC samples and obvious downregulation expression differences in tumor as nontumor specimens were found. In addition, hsa-miR-30d and hsa-let-7b were expressed in all tumor and nontumor samples (Figures 1 and 2). A significant downregulation of hsa-miR-30d was observed in NSCLC resected tissue, suggesting that hsa-miR-30d acts as a potential tumor-suppressor in NSCLC (Figure 2). Moreover, a significant expression alteration of hsa-miR-30d in stage III NSCLC tissues was found compared to stage I/II. The noticeable reduced expression of hsa-miR-30d was found in high-grade tumor, suggesting a dual role of the hsa-miR-30d in invasion and initiation of NSCLC.
Le et al have reported that hsa-miR-30d was overexpressed in pre-operative lung carcinoma patients and paired 10 day post-operative patients in whom its levels were significantly related with survival and its expression level showed that it could be a potentially novel non-invasive biomarker for diagnosis of lung cancer 22.
Our data are consistent with those of Markou et al’s 23, which revealed that expression levels of this miRNA were significantly downregulated in NSCLC tissues than in non-cancerous tissues. On the other hand, our results are in contrast with those in a study by Kobayashi et al 15, in which upregulation of hsa-miR-30d was shown in prostate cancer. In a previous study, hsa-miR-30d has been already shown to be downregulated in thyroid carcinogenesis via targeting polycomb protein enhancer of zeste 2 (EZH2) 24,25 and in squamous cell lung carcinoma 23. Moreover, hsa-miR-30d expression has revealed to be decreased in some malignancies including ovarian cancer cells via Transforming Growth Factor beta1 (TGF-beta1) and pancreatic cancer 25,26. Recently, hsa-miR-30d was found downregulated and it functions as a tumor suppressor by directly targeting the CCNE2 in NSCLC.
One reasonable description for the contradictory reports on miRNA expression in similar cancer types could be that they were the result of applying patients with different epigenetic alteration in genomic context with miscellaneous types, grades or stages of malignancy. Furthermore, has-mir30d has an especial seed sequence that has been demonstrated to bind to several critical genes which have essential role in Epithelial Mesenchymal Transition (EMT) as a key process for the initiation of cancer 26. Smoking status in patients with NSCLC is related with particular alterations to the epigenomic and genomic landscapes of lung cancer 27. Therefore, a noticeable underexpression level of hsa-miR-30d was seen in smokers of NSCLC, which is in agreement with those in a report written by Vucic et al 28.
Hsa-let-7b is one of the most commonly evaluated miRNA in human cancers, gastric cancer 29-31, melanomas 32 and lung cancer 33,34. As mentioned above, a significant upregulation of hsa-let-7b expression in NSCLC was found compared to that in their nontumor samples. Accordingly, ozcan et al reported that several members of let-7 cluster had a key role in colorectal carcinoma through Microtubule-associated Tumor Suppressor 1 (MTUS1) 35. Our findings are in accordance with those of Edmonds et al 36 which showed a significant increase of hsa-let-7b in stage I lung adenocarcinomas relapse patients and it appears to be involved in patients with NSCLC. Furthermore, Fassina et al have shown upregulation of hsa-let-7b in other let-7 families in adenocarcinoma in resected tumor specimens 33. In agreement with our findings, overexpression of hsa-let-7b in tissue of NSCLC patients is significantly correlated with type of NSCLC, suggesting that expression of hsa-let-7b might be useful as a prognostic marker. On the other hand, several studies showed downregulation of the let-7b in tumor cell lines 37 or tumor tissue when compared to their non-tumor 38.
It has been reported that overexpression of hsa-let-7b leads to decreased expression of Insulin-Like Growth Factor receptor 1 (IGF1R) at the post-transcriptional level introducing it as a tumor suppressor in multiple myeloma 39.
Differentially expressed hsa-let-7b was identified when smokers to nonsmokers were compared in NSCLC. Remarkably, the expression levels of both miRNAs were considerably lower in the tissue of the smoking NSCLC group than in the nonsmoking control group. The hsa-let-7b is a member of the let-7 family which regulates various cellular processes in cancers and is involved in the growth and proliferation through a potential role in angiogenesis 38.
The incongruence between these studies and ours led us to apply different methodologies and in addition, not to discriminate between different grades and stages of tumors. Furthermore, one of the possible reasons that can be explained is the conflicting role of let-7b, in which the stromal content of tumor microenvironment specimens such as Cancer-Associated Fibroblasts (CAF) can affect the consequence of expression results 40. One of the limitations of our study was the small sample size.
Our results provide evidence that miRNA expression distinguishes not only tumor tissue from normal tissue but also specific miRNAs for histopathological subtypes of NSCLC. Furthermore, using ROC curve analysis, has-hsa-miR-30d and hsa-let-7b were found to be potentially good tumor biomarkers to distinguish between NSCLC and non-cancer. Also, the expression level of hsa-miR-30d varied significantly in different stages. Remarkably, the pattern of expression in different stages was almost similar for hsa-let-7b-5p.
Conclusion :
Taken together, our data indicated significant expression alterations of hsa-miR-30d and hsa-let-7b in NSCLC patients, and that hsa-miR-30d could be used to discriminate between NSCLC and nontumor of lung cancer tissue specimens. Based on ROC analysis, it appeared that evaluation of both miRNAs is an informative diagnostic biomarker for lung malignancy. In addition, it seems that downregulations of hsa-miR-30d might contribute to tumor initiation and development. Our findings support the potential oncogenic role for has-let-7b-5p in NSCLC tumor development.
Acknowledgement :
We are grateful to Dr. Shams, the current head of the Baqiyatatllah Tumor Bank, Dr. Baghery, and Mrs. Chavoshi for their valuable help in supplying clinical specimens and providing patients’ clinicopathological information.
Conflict of Interest :
The authors declare that they have no competing interests.
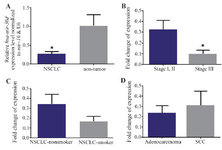
Figure 1. A) RT-qPCR analysis shows the mean values of relative hsa-miR-30d expression in Non-Small Cell Lung Cancer (NSCLC) and nontumor controls, with confidence intervals as the error bars. Note that the expression of hsa-miR-30d was significantly lower in 24 NSCLC tissues than that in the corresponding nontumor (p= 0.0382). Relative expression levels of hsa-miR-30d are demonstrated for different stages, B) and smoking status groups, C) of NSCLC patients. Note that the observed differences in expression were not statistically significant. D) A similar comparison in lung adenocarcinoma vs. Squamous Cell Carcinoma (SCC) samples. P-values of <0.05 were considered statistically significant.
|
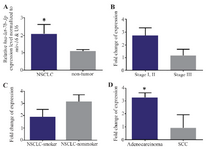
Figure 2. A) RT-qPCR analysis shows the mean values of relative hsa-let-7b expression in NSCLC and nontumor controls, with confidence intervals as the error bars. Note that the expression of hsa-let-7b was significantly upper in 24 NSCLC tissues vs. nontumor (p= 0.03). Relative expression alterations of hsa-let-7b are demonstrated for different stages, B) and smoking status groups, C) of NSCLC patients. D) A similar comparison in lung adenocarcinoma vs. SCC samples. P-values of <0.05 were considered statistically significant.
|
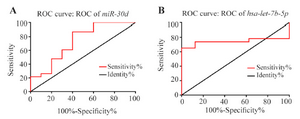
Figure 3. Receiver Operating Characteristic (ROC) curve analysis of the specificity and sensitivity of hsa-miR-30d (A) and hsa-let-7b (B) expression in discriminating between NSCLC and nontumor samples. The areas under the curve were 73 and 74%, respectively, which suggests that both miRNAs may be potentially tumor markers for NSCLC diagnosis. * Represents p<0.05.
|
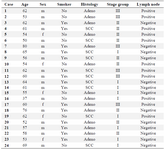
Table 1. Demographic and clinical features of surgically resected NSCLC patients
m: male, f: female. Smoking history; ≥20 pack years. SCC: small cell cancer; adeno; adenocarcinoma.
|

Table 2. Primer sequences used in RT-qPCR analysis
* V= G, A, C; N= G, A, T, C
|
|