Cloning and Expression of CD19, a Human B-Cell Marker in NIH-3T3 Cell Line
-
Abbasi-Kenarsari, Hajar
-
Immunology Research Center, Tabriz University of Medical Sciences, , Tabriz, Iran
-
Department of Immunology, School of Medicine, Tabriz University of Medical Sciences, Tabriz, Iran
-
Students Research Committee, Tabriz University of Medical Sciences, Tabriz, Iran
-
Shafaghat, Farzaneh
-
Department of Immunology, International Branch of Aras, Tabriz University of Medical Sciences, Tabriz, Iran
-
Baradaran, Behzad
-
Immunology Research Center, Tabriz University of Medical Sciences, Tabriz, Iran
-
Department of Immunology, School of Medicine, Tabriz University of Medical Sciences, Tabriz, Iran
-
Shanehbandi, Dariush
-
Immunology Research Center, Tabriz University of Medical Sciences, Tabriz, Iran
-
 Kazemi, Tohid
Department of Immunology, School of Medicine, Tabriz University of Medical Sciences, Tabriz, Iran, Tel: +98 411 3364665, Email: tohid_kazemi@yahoo.com
Kazemi, Tohid
Department of Immunology, School of Medicine, Tabriz University of Medical Sciences, Tabriz, Iran, Tel: +98 411 3364665, Email: tohid_kazemi@yahoo.com
-
Immunology Research Center, Tabriz University of Medical Sciences, Tabriz, Iran
-
Department of Immunology, School of Medicine, Tabriz University of Medical Sciences, Tabriz, Iran
Abstract: Background: CD19 is a pan B cell marker that is recognized as an attractive target for antibody-based therapy of B-cell disorders including autoimmune disease and hematological malignancies. The object of this study was to stably express the human CD19 antigen in the murine NIH-3T3 cell line aimed to be used as an immunogen in our future study.
Methods: Total RNA was extracted from Raji cells in which high expression of CD19 was confirmed by flow cytometry. Synthesized cDNA was used for CD19 gene amplification by conventional PCR method using Pfu DNA polymerase. PCR product was ligated to pGEM-T Easy vector and ligation mixture was transformed to DH5α competent bacteria. After blue/white selection, one positive white colony was subjected to plasmid extraction and direct sequencing. Then, CD19 cDNA was sub-cloned into pCMV6-Neo expression vector by double digestion using KpnI and HindIII enzymes. NIH-3T3 mouse fibroblast cell line was subsequently transfected by the construct using Jet-PEI transfection reagent. After 48 hours, surface expression of CD19 was confirmed by flow cytometry and stably transfected cells were selected by G418 antibiotic.
Results: Amplification of CD19 cDNA gave rise to 1701 bp amplicon confirmed by alignment to reference sequence in NCBI database. Flow cytometric analysis showed successful transient and stable expression of CD19 on NIH-3T3 cells (29 and 93%, respectively).
Conclusion: Stable cell surface expression of human CD19 antigen in a murine NIH-3T3 cell line may develop a proper immunogene which raises specific anti-CD19 antibody production in the mice immunized sera.
Introduction :
CD19 (95 kDa) is a type I transmembrane glycoprotein with 556 amino acids in length, belongs to immunoglobulin superfamily and also known as B4 1-3. CD19 antigen as a pan B cell marker is expressed by almost all B lymphocyte developmental stages from early pre-B cell until final differentiation to plasma cell, and also on follicular dendritic cell 4,5. CD19 gene contains 15 exons and located on short arm of chromosome 16 6,7. This molecule acts as B cell co-receptor in association with other surface B cell receptors including CD21, CD81 and CD225, and modulates signaling thresholds of BCR activation 8-10. CD19 represents as potential B cell target for therapeutic approaches in the treatment of B cell malignancies and autoimmune diseases 11-13. This marker is a reasonable candidate for antibody targeting since it is expressed on neoplastic cells in a variety of B cell leukemias and lymphomas 14,15. Furthermore, CD19 has a broad expression throughout the developmental stages on B lymphocytes but is not present on hematopoietic stem cells 16,17. Therefore, anti-CD19 monoclonal antibody could be useful in therapeutic and also diagnostic areas.
First described by Georges Kohler and Cesar Milstein 18, hybridoma technology is the most common way to produce mouse monoclonal antibodies. The first step, immunization, needs some important considerations. Among them, preparing proper immunogen plays pivotal role in obtaining high quality monoclonal antibodies. Owing to lack of glycosylation in prokaryotes 19, using recombinant proteins produced in eukaryotic instead of prokaryotic expression systems has some advantages e.g. restoring all potential epitopes. Stable expression of protein of interest in mouse cell line including NIH-3T3, on the other hand, bypasses the need for purification of recombinant protein though some technical ambiguities may be encountered. The other advantage of immunization of mouse by stably transfected cell line is intraperitoneal administration of the cells without adjuvant. Intradermal injection of purified immunogen mixed by an adjuvant may result in tissue destruction and side effects which weaken the animal 20.
Therefore, in this study, an attempt was made to clone the cDNA coding for CD19 molecule from Raji cell line for subsequent expression in mouse NIH-3T3 cell line and production of monoclonal antibodies against the extracellular part of molecule.
Materials and Methods :
Cell lines and bacteria: Raji and NIH-3T3 cell lines were purchased from National Cell Bank of Iran (NCBI, Tehran, Iran) and cultured in RPMI 1640 medium with L-glutamine (Gibco, Darmstadt, Germany) supplemented with 10% Fetal Bovine Serum (FBS) (Gibco, Darmstadt, Germany), 100 mg/ml Penicillin and 100 IU/ml Streptomycin (Gibco, Darmstadt, Germany) and incubated at 37ºC in a humidified incubator with 5% CO2. E. Coli strain DH5α (Promega, WI, USA) was used in cloning and grown in Luria Bertani medium.
Flow cytometry: To evaluate the surface expression of CD19 molecule on Raji cell line, the cells were subjected to indirect staining. First, 5×105 cells were washed by FACS washing solution (PBS 1× containing 0.1% NaN3). Then the cells were stained with mouse anti-human CD19 monoclonal antibody (Biolegend, London, UK) to a final concentration of 5 μg/ml. In the other tube, Raji cells were stained with isotype control antibody, mouse IgG1/κ with unrelated specificity (Biolegend, London, UK), as negative control for staining. Cells were incubated for 1 hr at 4°C and after two times of washing, resuspended in buffer containing secondary antibody, FITC-conjugated sheep anti-mouse immunoglobulin (Avicenna Research Institute, Tehran, Iran) at a 1/50 dilution. Incubation in a dark place for 1 hr at 4°C was followed by two washing steps. Finally, cells were resuspended in FACS washing solution and scanned in the flow cytometer (BD FACSCalibur flow cytometer).
RNA extraction and cDNA synthesis: 5×106 Raji cells were harvested and washed two times by RPMI 1640 culture medium. Cells were lysed with 1 ml of RNX-plus solution (CinnaGen, Tehran, Iran) and RNA extraction was done following the manufacturer's instructions. Briefly, 200 μl chloroform was added to cell lysate and incubated for 5 min on ice.
Following centrifugation and collection of the aqueous phase in new tube, the total RNA was precipitated by isopropanol in equal volume after incubation for 15 min. The RNA pellet was washed by 75% ethanol and resolved in RNase-free double distilled water. Prior to cDNA synthesis, RNA concentration was determined by UV spectrophotometry and integrity of extracted RNA was checked by agarose gel electrophoresis. 5 μg of total RNA together with 1 μl random hexamer (N6) primer (Thermo Fisher Scientific, Inc., MA, USA) were heated at 65°C for 5 min and placed on ice. Then, 200 U MMuLV reverse transcriptase enzyme (Thermo Fisher Scientific, Inc., MA, USA), 4 μl of 5× reaction buffer (Thermo Fisher Scientific, Inc., MA, USA) and 1 μl of 10 mM dNTP (Thermo Fisher Scientific, Inc., MA, USA) were added to the reaction and incubated at 25°C for 10 min followed by 60 min at 42°C. To terminate the cDNA synthesis, the reaction was heated at 70°C for 10 min. Finally, β-actin as a housekeeping gene was amplified for confirming synthesis of cDNA.
Primer design and polymerase chain reaction: Reference sequence of human CD19 mRNA isoforms available in NCBI database (variant 1: NM_001178098.1, variant 2: NM_001770.5) were used to design gene-specific primers for PCR amplification of cDNA. Variant 2 uses an alternate in-frame splice site in the 3' coding region, compared to variant 1 that results in a shorter protein (isoform 2), compared to isoform 1. In order to sub-clone the cDNA in pCMV6-Neo expression vector, KpnI and HindIII restriction sites were embedded at the proximity of start codon of forward primer and 3´end of stop codon in reverse primer, respectively. Also, Kozak sequence (GCCACC) was added to forward primer after the sequence for KpnI restriction site and before ATG codon for efficient translation in eukaryotic expression system. Forward and reverse primers for specific amplification of CD19 cDNA were designed as following: forward primer: 5´-GGTACCGCCACCATGGCACC TCCTCGCCTCCTCTTC -3´ and reverse primer: 5´- AAGCTTGCCACCTGAGGATCACCTGGTGC-3´.
To optimize PCR conditions, different concentrations of MgCl2 and a range of annealing temperatures were examined using Taq DNA polymerase (CinnaGene, Tehran, Iran). For amplification of CD19 cDNA, PCR was performed in a 25 μl reaction solution contained 2.5 μl 10×PCR buffer, 1.5 μl 10 mM dNTPs, 1 μl of each primer (10 pmol/μl), 0.5 μl Pfu DNA polymerase (10 U/μl) (Thermo Fisher Scientific, Inc., MA, USA), 1 mM MgSO4 and 1 μl cDNA. Each PCR reaction underwent initial denaturation at 95°C for 5 min followed by 39 cycles of denaturation (95°C for 30 s), annealing (different temperatures for 30 s) and extension (72°C for 2.5 min), and final extension at 72°C for 10 min. For proper TA cloning, A-tailing was done at 72°C for 7 min using Taq DNA polymerase. PCR products were subjected to agarose gel electrophoresis, visualized by ethidium bromide at final concentration of 5 μg/ml and documented in UVP Gel Documentation System (UVP, CA, USA).
TA-cloning of CD19 cDNA: PCR product was extracted from agarose gel using GeneJet Gel Extraction Kit (Thermo Fisher Scientific, Inc., MA, USA) and then cloned into pGEM-T Easy vector (Thermo Fisher Scientific, Inc., MA, USA). Ligation was performed by 3 units of T4 DNA ligase (Thermo Fisher Scientific, Inc., MA, USA), followed by overnight incubation at 4°C. Then ligation mixture was transformed into DH5α competent bacteria by heat shock method. Briefly, 10 ml of ligation mixture was added to 100 ml of bacteria and after gentle mixing, incubated for 20 min on ice. After that, the mixture was subjected to 42°C for 1 min and then chilled on ice for 3 min. For refreshing transformed bacteria, LB broth medium was added to cells and incubated for 1 hr at 37°C with shaking at 150 rpm. Then transformed bacterial cells were plated onto LB agar containing 100 µg/ml Ampicillin (Dana, Tabriz, Iran), 40 µl 20 μg/ml X-Gal (Thermo Fisher Scientific, Inc., MA, USA) and 40 µl 0.1 mM IPTG (Thermo Fisher Scientific, Inc., MA, USA). After overnight incubating at 37ºC, colony-PCR using gene-specific primers and conditions similar to PCR reaction for amplification of CD19 cDNA were applied on white colonies. Then single colony containing desired insert was cultured in LB broth containing Ampicillin and incubated in shaker incubator (250 rpm) at 37°C overnight for miniprep preparation using Gene JET Plasmid miniprep kit (Thermo Fisher Scientific, Inc., MA, USA) according to the manufacturer’s recommendations. Next, pGEM-T Easy/CD19 gene construct was sequenced using T7 promoter and SP6 universal primers.
Sub-cloning in pCMV6-Neo expression vector: pGEM-T Easy/CD19 gene construct was double digested using 10 units of each KpnI (as partial digestion for 5 min) and HindIII (as complete digestion for 16 hr) restriction enzymes (Thermo Fisher Scientific, Inc., MA, USA). Digested CD19 cDNA was sub-cloned into pCMV6-Neo expression vector digested by the same enzymes. Then pCMV6-Neo/CD19 construct was transformed into DH5α competent bacteria and selected by plating on LB agar containing Ampicillin. Existence of gene in recombinant vector was verified by colony-PCR using gene-specific primers on some of randomly selected colonies and also, by double digestion of prepared miniprep from positive colony. Ultimately, to confirm sequence fidelity and insert orientation, pCMV6-Neo/CD19 construct was sequenced using vector-specific primers, V1.5 and XL39.
Eukaryotic expression of human CD19 in NIH-3T3: 105 NIH-3T3 cells were cultured overnight in 6-well cell culture plate and transfected by the mixture of 3 mg pCMV6-Neo/CD19 recombinant construct and 6 ml JetPEI transfection reagent (Polyplus-transfection Inc. NY, USA) according to manufacturer’s recommendations. In parallel, cells were transfected by expression vector devoid of CD19 cDNA (mock transfection). 48 hr after transfection, transient expression of CD19 antigen was examined by flow cytometry methods, as described earlier. To obtain stable expression, cells were cultured in complete cell culture medium containing G418. Starting concentration was determined by titration of G418 antibiotic on NIH-3T3 cells. Growing of transfected cells and dying of untransfected cells were monitored and live cells were subjected to gradually increasing concentrations of G418 up to 1500 mg/ml during two months. Stable expression was confirmed by flow cytometry method as described earlier.
Results :
Surface expression of CD19: Flow cytometric analysis of Raji cells, as the source for amplification of CD19 cDNA and positive control for surface expression of CD19 in further experiments, showed positive reactivity with anti-CD19 monoclonal antibody (M2: 98%) in comparison with cells stained by isotype control (M1: 2%). On the other hand, expression of CD19 mRNA was successfully showed by amplification of CD19 cDNA in PCR method (Figure 1A), as described later.
Amplifying and TA-cloning of CD19 cDNA: Initial optimization using Taq DNA polymerase showed specific band in annealing temperature of 64ºC and 1 mM concentration of MgCl2. Subsequent PCR reaction using Pfu DNA polymerase led to a single band with expected size (1701 bp) (Figure 1A). PCR product was ligated into pGEM-T Easy vector by TA-cloning method and then transformed to competent DH5α bacteria using heat shock method. Blue-white selection was used for screening white colonies containing recombinant plasmid. Among them, eight white colonies were subjected to colony-PCR with gene-specific primers and two of them showed the expected band (Figure 1B). Then, miniprep was prepared from single positive colony and double digested with KpnI and HindIII restriction enzymes. Because of existence of one restriction site for KpnI in CD19 gene at nucleotide position 603, pGEM-T Easy-CD19 construct was subjected to partial digestion by KpnI and complete digestion by HindIII and led to excision of insert with length of 1701 bp (Figure 1C). Alignment of sequencing result with reference sequence in NCBI database showed complete matching with CD19 mRNA isoform 2 except at nucleotide position 4 where C was replaced with G in order to improve the expression of gene (Figure 2).
Construction of recombinant pCMV6-Neo/CD19 construction: pGEMT-easy/CD19 recombinant cloning vector was digested by KpnI and HindIII restriction enzymes. Excised CD19 cDNA with 1701 bp band was ligated into double digested pCMV6-Neo expression vector by the same enzymes and pCMV6-Neo/CD19 construction was transformed into competent DH5α bacteria. After transformation, eight colonies were obtained from cultured DH5α bacteria in LB agar/ampicillin medium. After performing colony-PCR reaction, three colonies showed successful insertion and one of the colonies was selected for preparation of miniprep, direct sequencing and preparation of maxiprep for subsequent experiments.
Transfection of NIH-3T3 cells and expression of human CD19 protein: NIH-3T3 mouse fibroblastic cell line was transfected by pCMV6-Neo/CD19 recombinant expression vector using JetPEI transfection reagent and transient expression of CD19 was confirmed by surface flow cytometry 48 hr after transfection (Figure 3A). After that, pCMV6-Neo/CD19- and mock-transfected and also untransfected NIH-3T3 cells were subjected to growth in complete cell culture medium containing 400 mg/ml G418 which was the minimal dose for elimination of untransfected NIH-3T3 cells. After 3 days, untransfected cells died and transfected cells continued to grow in G418 up to 1500 mg/ml during two months. Cells were finally analyzed by surface flow cytometry and significant positive reactivity of pCMV6-Neo/CD19- transfected cells was observed (93%) (Figure 3B). On the other hand, untransfected and mock-transfected NIH-3T3 cells showed no positive signal in flow cytometry (4 and 6%, respectively) (Figures 3C and D).
Discussion :
cDNA coding for human CD19 was first cloned and described by Tedder et al from human tonsil cDNA library 3. Located on chromosome 16, CD19 gene codes for a transmembrane protein with exclusive expression in B cell lineage 4-6. CD19 as a promising target for immunotherapy of B cell malignancies and autoimmune diseases has been the focus of interest by several investigators. Several clinical trials have been conducted to target this marker using various formats, including bispecific antibodies, Fc-engineered antibodies, antibody-drug conjugates (ADCs) and anti-CD19 chimeric antigen receptors (CARs) for therapeutic approaches 11-13, 21. The objective of this study was cloning of cDNA coding for human CD19 and stable expression in NIH-3T3 mouse fibroblast cell line to produce proper immunogen for further experiments including production of CD19-specific monoclonal antibodies. cDNA cloned in this study was amplified from cDNA synthesized using total RNA of Raji Burkitt’s lymphoma cell line with high expression of CD19 22 by specifically designed primers. Comparing the sequence of cloned cDNA to NCBI database showed successful alignment to short isoform (isoform 2) of human CD19. This isoform lacks amino acid Alanine at position 496 compared to long isoform (isoform 1). Isoform 2 is the canonical isoform of human CD19 and predominantly expressed by B-cell lineage. Verified cDNA of full length human CD19 sub-cloned in eukaryotic expression vector and resulting expression construct transfected in NIH-3T3 cells. Results of flow cytometry analysis showed successful stable expression of CD19 on the surface of NIH-3T3 cell line.
According to modified protocol introduced in 1970s for producing mouse monoclonal antibodies 23, the first and one of the most important steps of hybridoma technology is preparing a good and proper immunogen. It should be intact as much as possible for preserving all potential epitopes. On the other hand, using full-length protein with proper post transcriptional modifications similar to that occurs for native protein, is helpful for obtaining functionally more active and useful mAbs 24. Additionally, immunization of animal with purified protein is done by mixing antigen with Complete (CFA) and Incomplete Freund’s Adjuvant (IFA) usually administered subcutaneously. According to American Association for Laboratory Animal Science (IACUC) Policy on Administering Complete Freund’s Adjuvant 20, this protocol could induce unwanted local inflammation causing skin ulcerations and draining sinuses with granulomas and may lead to improper antibody response to antigen and limited number of antigen-specific clones. All above mentioned reasons force us to produce recombinant proteins in eukaryotic expression systems and among them purification step is tedious and challenging. However, immunization of mouse using stably-transfected murine NIH-3T3 cell line needs neither adjuvant nor purification and the route of injection is intraperitoneal with minimal side-effects for the animal. Stable expression of cDNA coding for protein of interest in mouse fibroblast cell line, NIH-3T3, has been previously used by our group as an approach to prepare immunogen for immunization of mouse 25.
Conclusion :
Due to mouse origin of NIH-3T3 cell line, only the ectopically expressed protein would be immunogen in immunized mouse and stable CD19-expressing NIH-3T3 cells could be used as a proper immunogen in subsequent studies for production of diagnostic and therapeutic monoclonal antibodies using hybridoma technology.
Acknowledgement :
This study was supported by grant from Immunology Research Center, Tabriz University of Medical Sciences.
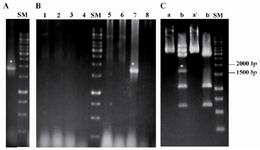
Figure 1. Cloning and sub-cloning of CD19 cDNA. A) Amplification of specific band for human CD19 cDNA using Pfu DNA polymerase; B) Colony-PCR reaction on eight white colonies (1-8) after blue/ white selection. C) Excision of 1701 bp band for human CD19 cDNA after double digestion of the construct using KpnI and HindIII restriction enzymes. Lanes (a) and (a'): undigested pGEMT-easy/CD19 construct, Lanes (b) partial and complete digestion by KpnI and Hind III, respectively and (b'): complete digestion by both KpnI and HindIII. SM: DNA size marker (bp). Asterisks (*) point the desired band.
|

Figure 2. Alignment of amplified cDNA for canonical isoform of human CD19 reference sequence in NCBI database. Comparing the 1701 bp amplified sequence with reference sequence for short isoform (variant 2) of human CD19 showed complete alignment. Only 5` and 3` of alignment has been briefly showed.
|

Figure 3. Flow cytometric analysis of expression of human CD19. Using specific monoclonal antibody, expression of human CD19 for transient (A) and stable expression (B); untransfected NIH-3T3 (C); mock-transfected NIH-3T3 (D); and Raji cell lines (E, as positive control) were analyzed.
|
|