Induction of Strong and Specific Humoral and T-helper 1 Cellular Responses by HBsAg Entrapped in the Methanobrevibacter smithii Archaeosomes
-
Delbaz, Seyed Ali
-
Department of Hepatitis and AIDS, Pasteur Institute of Iran, Tehran, Iran
-
Sadat, Seyed Mehdi
-
Department of Hepatitis and AIDS, Pasteur Institute of Iran, Tehran, Iran
-
Rahimi, Pooneh
-
Department of Hepatitis and AIDS, Pasteur Institute of Iran, Tehran, Iran
-
Bolhassani, Azam
-
Department of Hepatitis and AIDS, Pasteur Institute of Iran, Tehran, Iran
-
Bahramali, Golnaz
-
Department of Hepatitis and AIDS, Pasteur Institute of Iran, Tehran, Iran
-
Motevalli, Fateme
-
Department of Hepatitis and AIDS, Pasteur Institute of Iran, Tehran, Iran
-
Davari, Mehdi
-
Department of Hepatitis and AIDS, Pasteur Institute of Iran, Tehran, Iran
-
Vakily, Habib
-
Department of Hepatitis and AIDS, Pasteur Institute of Iran, Tehran, Iran
-
 Sharifat Salmani, Ali
Department of Biology, Science and Research Branch, Islamic Azad University, Tehran, Iran, E-mail: alisharifat@gmail.com
Sharifat Salmani, Ali
Department of Biology, Science and Research Branch, Islamic Azad University, Tehran, Iran, E-mail: alisharifat@gmail.com
-
Department of Biology, Science and Research Branch, Islamic Azad University, Tehran, Iran
-
Borhan Nobari, Maryam
Department of Hepatitis and AIDS, Pasteur Institute of Iran, Tehran, Iran, Tel: +98 66953311; Email: azad237@yahoo.com
-
Department of Hepatitis and AIDS, Pasteur Institute of Iran, Tehran, Iran
Abstract: Background: Application of adjuvants with microbial origins is a recently highlighted approach in the vaccinology trials. Archaeosomes are among these microbial compounds with both adjuvant and liposomal activities and features. Methods: In the present study, recombinant HBsAg encapsulated into Methanobrevibacter smithii (M. smithii) archaeosomes. Balb/c mice immunized with this compound and humoral and cytokine secretion pattern of immunized models analyzed. Results: Frequency of IFN-γ secreting cells in the HBsAg-containing archaeosomes group was significantly higher than HBsAg and HBsAg+C/IFA groups (p≤0.05). IgG2a titer in the sera of HBsAg-containing archaeosomes group was also significantly higher than this subclass titer in the other groups (p≤0.05). Conclusion: Analysis of induced responses revealed the Immunopotentiating characteristics of M. smithii archaeosomes in the induction of T-helper 1 responses according to the dominance of IgG2a subtype and IFN-γ secreting splenocytes of immunized mice.
Introduction :
Although available recombinant Hepatitis B vaccines are efficiently applied in the prevention of Hepatitis B virus (HBV) infection, but no therapeutically efficacious vaccine is introduced for the clearance of HBV infection in the carriers and chronically infected patients. An ideal HBV vaccine would be both preventive and therapeutic which means that
this vaccine should promote the production of specific neutralizing antibodies and cell-mediated responses (particularly T helper 1 and cytotoxic T-cells) against the virus. One of the main obstacles in the procedure of such a vaccine production is the lack of suitable adjuvants with cellular responses inducing features. The classical adjuvant for vaccines, and until recently the only FDA-approved adjuvant, is alum which is used in the commercially available HBV vaccines 1. Alum provides a particle upon which the vaccine is precipitated. Although precipitation onto alum promotes uptake of the immunogen, alum is a poor activator of DCs and does not induce the production of IL-12 2. As a result, vaccines containing alum initiate a Th2-type antibody response 3.
Complete Freund’s adjuvant (CFA) is another powerful adjuvant consisting of inactivated Bacillus Calmette-Guerin (BCG) and a mixture of different TLR ligands in a mineral oil solution 1. Although BCG is a strong activator of DCs and induces a Th1 response, CFA is not approved for human use 1. Application of adjuvants with microbial origins is a recently highlighted approach in the vaccinology trials 4. Archaeosomes are among these microbial compounds with both adjuvant and liposomal activities and features 3. Archaeosomes are liposomes made of polar lipids obtained from archaeal (archaebacterial) cells. The mammalian-like archaeal lipids consist of isoprenoid chain glycerolipids. Archaeol (di-O-phytanylglycerol) is double ether of sn-1-glycerol where positions 2 and 3 are bound to phytanyl residues. The archaeols are Archaea homologs of Diacylglycerols (DAGs) 5. In certain archaea such as Methanobrevibacter smithii (M. Smithii), archaeol is found in the dimer form namely caldarchaeol which enhances the stability of archaeosomes originated from this archaea 6.
Some immunologically important features of archaeosomes include high stability of archaeal lipids as building blocks of archaeosomes which leads to long-lasting memory of immune responses, immunopotent interaction with Antigen Presenting Cells (APC) and most importantly shifting and leading the antigen processing pathway to MHC class I mechanisms and subsequently intensifying CD8+ T-cell responses 3,5. Available vaccines are mostly designed according to the immunostimulatory properties of Hepatitis B surface Antigen (HBsAg) which is also called envelope antigen. The HBV genome encodes for three related envelope proteins termed L, M, and S. The three envelope proteins are produced from a single Open Reading Frame (ORF) through alternative translation start sites. HBsAg is an antigenically heterogeneous protein, with a common antigen designated a, and two pairs of mutually exclusive antigens, d and y, and w and r, resulting in 4 major subtypes, namely adw, ayw, adr and ayr 7.
In the present study, recombinant HBsAg (Recombinant HBV HBsAg Antigen, Subtype adw, Acrobiosystems, USA) was encapsulated into M. smithii archaeosomes. Balb/c mice were immunized with this compound and humoral and cytokine secretion patterns of immunized models were analyzed.
Materials and Methods :
Preparation of archaeosomes: M. smithii (DSMZ 2375) was grown in fermenter (Nova-Paljas, Contact-flow., Netherland) containing DSMZ recommended medium (Table 1) for 72 hr in anaerobic atmosphere (80% H2, 20% CO2). Archaeal cells were harvested by centrifugation at 3800 g for an hour and kept at -20°C to become completely frozen and then thawed at room temperature. Archaeal lipids were extracted and antigen free and antigen containing archaeosomes were prepared as previously described 6.
Immunization: Five animal groups each consisted of seven pathogen-free, female BALB/c mice (6-8 weeks of age-average 20 g of weight) were handled according to the international animal care ethics and immunized with HBsAg, HBsAg-containing archaeosomes (HBsAg+ Arch), HBsAg-free archaeosomes (Arch) and HBsAg with complete/incomplete Freund’s adjuvant (HBsAg+C/IFA). Mice were immunized subcutaneously at the base of the tail with 2 μg HBsAg, archaeosome-HBsAg (2 μg of HBsAg in 0.5 mg of lipid/100 μl of PBS), 0.5 mg of HBsAg-free archaeosomes and 2 μg HBsAg formulated in C/IFA; two booster immunizations were carried out three and six weeks after the first immunization. Immunization with Freund’s adjuvant was carried out as the usual protocol (the first immunization with complete and second and third immunizations with incomplete Freund’s adjuvant). Phosphate Buffered Saline (PBS) was injected to another group as the negative control.
Immunological analysis: Total and subclass titer on HBsAb: Anti HBsAg humoral response was assessed by ELISA in different immunization groups. Wells of microtiter plates were coated with 1 μg.ml-1 recombinant HBsAg (Acrobiosystems, USA) and blocked by bovine serum albumin. 150 μl of diluted sera of immunized mice was added to each well. Sera were 1:50 and 1:100 diluted for total IgG and related subtypes measurements, respectively. Dilution rates were determined by pre-testing serially diluted pooled sera of test groups against the coated antigens. HRP-conjugated anti mouse total and subtype IgG (Thermo Fisher Scientific Inc, USA) was added to detect the specific HBsAb IgG molecules 4.
Cytokine assay: ELISpot and ELISA: Frequency of IFN-γ and IL-4-secreting splenocytes of immunized models was determined by ELISpot assay (e-bioscience, CA) two weeks after the last immunization. The concentrations of both cytokines in the splenocytes culture medium were also assayed by ELISA (UcyTech, Netherlands). Single-cell cultures of spleen cells (105 cells/well for ELISpot and 106 cells/ml for ELISA) were prepared in the presence of 10 M HBsAg for 40 and 72 hr at 37°C, respectively. The applied medium was complete RPMI-1640 supplemented with 10% FBS, 2 mM L-glutamine, 100 U/ml penicillin and 100 μg/ml streptomycin. The wells of ELISpot assay were coated by anti-mouse IFN-γ and IL-4 and prepared by the manufacturer. The secondary antibody was biotinylated. HRP-labeled streptavidin and the substrate of HRP were finally added. The Spot Forming Cells (SFCs) were developed and counted in the ELISpot assay using stereo microscope as the frequency index of IL-4 and IFN-γ secreting splenocytes. The results were expressed as the number of SFCs per 106 input cells. The specificity of cytokine secretion was controlled by the frequency of SFCs in the presence of an irrelevant peptide (aa 132-145 HCV-Core) and phytohemagglutinin (PHA) was applied as the positive control 8. ELISA was performed according to the procedure recommended by the manufacturer. The sensitivity limit of ELISA kit was 10 pgml-1 for both cytokines.
Statistical method: All experiments were carried out in triplicate and repeated three times. Differences between results and groups were statistically analyzed by Mann-Whitney non-parametric test and one way ANOVA (ANALYSE-IT 2.20 software). The p-values less than 0.05 were considered significant.
Results :
Immune responses: Total and subclass IgG titer: As it was previously mentioned, total anti HBs IgG and related subclasses in the sera of immunized mice were determined by ELISA method. Total IgG titer in the sera of mice immunized with all of the HBsAg containing formulations was significantly higher than the group immunized with HBsAg-free archaeosomes and negative control group. The highest titer was detected in the sera of HBsAg-containing archaeosome immunized group (Figure 1). Total anti HBs IgG in HBsAg-containing archaeosomes group was even significantly higher than the group immunized with HBsAg+C/ICFA (p≤0.05). Immunization with HBsAg free archaeosomes did not elicit significant HBsAb in comparison to the negative control group. IgG subclasses titers were also analyzed to estimate the orientation of cell-mediated responses. IgG2a was the dominant subclass in the group immunized with HBsAg-containing archaeosomes while this dominance was not observed in other immunization groups. IgG2a titer in the sera of HBsAg-containing archaeosomes group was also significantly higher than this subclass titer in other groups (p≤0.05). The ratio of IgG2a to total IgG in this group was 0.71 while the ratio of IgG2b and IgG1 to total IgG was 0.32 and 0.33, respectively.
Although IgG2b and IgG1 subclasses were both efficiently elicited in HBsAg, HBsAg-containing archaeosomes and HBsAg+C/IFA immunization groups, but there was no significant difference between the titer of these subclasses in the above-mentioned groups.
Cytokine assay: ELISA determined totally different patterns of IFN-γ and IL-4 concentration in the culture media of splenocytes. The concentration of IFN-γ was significantly higher in the splenocytes culture medium of mice immunized by HBsAg-containing archaeosomes in comparison to the group immunized by HBsAg+C/IFA (p≤0.05) (Figure 2). Significant difference was not shown in the IL-4 concentration secreted by splenocytes of mice immunized by HBsAg containing archaeosomes and HBsAg+C/IFA.
Splenocytes of mice immunized with HBsAg, HBsAg-containing archaeosomes and HBsAg+C/IFA specifically produced IFN-γ and IL-4 since the frequency of both cytokines secreting cells was insignificant in the presence of irrelevant peptide (HCV core) in the ELISpot assay (Figure 3). Frequency of IFN-γ secreting cells in the HBsAg-containing archaeosomes group was significantly higher than HBsAg and HBsAg+C/IFA groups (p≤0.05) while the frequency of these splenocytes in HBsAg and HBsAg+C/IFA immunized groups didn’t show a significant difference. In comparison with the group immunized by antigen-free archaeosomes, frequency of IL-4 secreting splenocytes was also remarkable in the main immunization groups particularly in the group immunized with HBsAg+C/IFA. Ratio of IFN-γ/IL-4 SFCs in the HBsAg-containing archaeosomes group was higher than the other groups (HBsAg-containing archaeosomes: 1.94, HBsAg: 1.34, HBsAg+C/IFA: 0.74). Among the groups immunized with the HBsAg-containing formulations, significantly more frequent IL-4 SFCs was observed only in the HBsAg+C/IFA immunization group.
Discussion :
HBV commercial vaccines mostly promote efficient humoral responses for prevention of infection. A considerable concern in HBV vaccine researches is the lack of a therapeutic vaccine for clearance of infection through the augmentation of T helper 1 and CD8+ T-cells 9. Lack of an applicable adjuvant to be formulated in the therapeutic HBV vaccine candidates has been a consistent problem in vaccine researches. Most of known adjuvants are not desirable for human use because of undesirable reactions or their weak effect on the induction of cellular responses 10. Archaeal liposomes (archaeosomes) are among newly introduced adjuvants with potency to augment humoral and cell-mediated responses, particularly immune responses with the dominance of T-helper 1 and CD8+ T-cells 11.
In the present study, an attempt was made to evaluate adjuvant properties of M. smithii archaeosomes to induce humoral and cellular responses by the assessment of IgG (total and subclass) and cytokine responses, respectively. Total anti HBs IgG and related subclasses were analyzed by ELISA and all indicated the potency of HBsAg-containing archaeosomes to induce significant IgG responses against HBsAg in comparison to other groups immunized with HBsAg and HBsAg+C/IFA (p<0.05). The advantage of archaeosomes to induce strong humoral responses to entrapped antigen has been previously confirmed for varied antigens (hen egg lysozyme, ovalbumin, cholera toxin) by various immunization routes 3,12. IgG subclasses analysis verified the dominance of IgG2a in comparison to IgG1 in the HBsAg-containing archaeosome immunized mice as a sign of T-helper 1 orientation of cell-mediated responses since supremacy of IgG2a and IgG1 is commonly considered as the dominance of T-helper 1 and T-helper 2 sub-population responses, respectively 13. The higher ratio of IgG2a/total IgG titer (0.71) in comparison with the ratio of IgG1/total IgG titer (0.33) in the HBsAg-containing archaeosome immunized mice sera is another sign of supremacy of T-helper 1 responses. Dominance of T-helper 1 oriented responses against viral infections would be a promising achievement in the application of archaeal adjuvants in the development of a therapeutic HBV vaccine.
The antibody secretion pattern was verified by the ELISpot results since the frequency of IFN-γ secreting splenocytes was significantly higher than IL-4 SFCs following the immunization with HBsAg-containing archaeosomes. IFN-γ secreting splenocytes in the group immunized by HBsAg-containing archaeosomes were also significantly higher in comparison to the groups immunized by HBsAg and HBsAg formulated in Freund’s adjuvant (p<0.05).
Similar to the results of IgG subclasses analysis that indicated the ratio of IgG2b/total IgG titer as the highest ratio between all subclasses, the ratio of IFN-γ/IL-4 SFCs was also the highest one in the group immunized by HBsAg-archaeal adjuvant as an indication of T-helper 1 oriented responses. According to the ability of M. smithii archaeosomes in the enhancement of MHC class I antigen presentation and the supremacy of IgG2a subtype and IFN-γ in the mentioned group, CTLs and T-helper 1 cells responses were suggested as the cellular responses induced by this administered formulation 14.
Although Hepatitis B vaccine is not a vague concept now, at least in the prevention of infection, findings of the present study would indicate the potency of M. smithii archaeosomes to induce strong T-helper 1 and IFN-γ mediated responses against HBsAg as a model of a viral antigen. However, there is also a clinical need for a therapeutically effective vaccine for Hepatitis B that induces robust cell-mediated response capable of viral clearance in the chronic infection. There are evidences indicating the impairment of T-helper 1 responses during the HBV infection leading to viral immune escape and consequently chronic HBV infection 15. The immunopotency of HBsAg-containing archaeosomes in the induction of T-helper 1 responses and IFN-γ production would be also therapeutically efficient to compensate T-helper 1 cells deficiency during HBV infection. Probable mechanism for enhancement of cellular immunity by this formulation is related to augmentation of immunological interactions and recruitment of antigens via macrophages and other APC 11.
Other efforts for development of a therapeutic HBV vaccine have been made previously. Chuai et al evaluated the effect of several adjuvants including alum, CpG and polyriboinosinic polyribocytidylic acid [poly(I:C)] to enhance anti-HBV response when boosted with recombinant adenoviral vector vaccine. Among the adjuvants tested, CpG/alum and poly(I:C)/alum combinations induced the specific production of high antibody titres with a Th1bias. In comparison to the present study, the frequency of IFN-γ SFCs following their immunization was considerably less than the frequency induced by HBsAg-containing archaeosome immunization. D. Morrey et al applied a Cationic Lipid DNA Complex (CLDC) as an adjuvant for HBsAg. Their combination efficiently induced both B and T-cells responses and they suggested that CLDC can be a promising adjuvant for therapeutic HBV vaccines although the orientation of the cellular responses was not specified in their study since IFN-γ was the only assayed cytokine.
In contrast, the present study assayed IFN-γ and IL-4 as an index for T helper 1 and T helper 2 responses, respectively. Buchmann et al evaluated the potential of a formulation comprising particulate HBsAg and core antigen (HBcAg), and the saponin-based ISCOMATRIX™ adjuvant for its ability to stimulate T and B cell responses in C57BL/6 mice. Their candidate vaccine induced strong CD8+ T-cell responses and efficiently reduced HBcAg expression in the liver of immunized mice. Application of archaeosomes in combination with different antigens is a vastly studied concept in vaccine researches. Priming and boosting with bovine serum albumin (BSA) entrapped in archaeosomes were shown to induce significantly higher antibody titers in comparison to immunization with the same antigen entrapped in conventional liposomes 16.
The advantage of archaeosomes to induce strong humoral responses to entrapped antigen has been confirmed for various antigens (hen egg lysozyme, ovalbumin, cholera toxin), and by multiple immunization routes (IP, IM, SC) and in murine strains of different genetic background (C3H/HeJ, C57BL/6, BALB/c). Similar to the results of the current study, analyzing the antibody isotype distribution in mice immunized with antigen in varied archaeosomes, indicated induction of strong IgG1, IgG2a and IgG2b titres, in contrast to alum that evokes little IgG2a isotype switching 3. Archaeosomes originated from certain archaea such as M. smithii and Thermoplasma acidophilum are more stable due to high percentage of caldarchaeols in their structures 17. This type of archaeosomes limits phagolysosomal antigen processing and enhances MHC class I presentation 18. This would be potentially beneficial in the induction of MHC class I and CTL responses against intracellular and viral infections. These responses are particularly favorable in the clearance of infected hepatocytes during HBV infection 19.
Conclusion :
According to the importance of cellular responses along with specific antibody reactions in all vaccinology aspects, the characteristic of elicited responses is not only applicable in HBV vaccine researches but also would be considered in other vaccine researches as well.
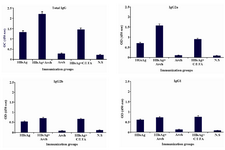
Figure 1. Total and subclass IgG response elicited by different immunization formulations. Each formulation is abbreviated on the horizontal axis of diagrams (see text for detailed materials and methods). Data show the obtained ELISA results for optimum dilution of mice sera against the coated HBsAg. Total IgG was determined at 1:50 dilution of mice sera, isotype-specific antibodies IgG2a, IgG2b and IgG1 were determined at 1:100 dilution of mice sera. Optimum dilutions were determined prior to the comparisons, by testing serially diluted sera pooled from individual mice of test groups against the coated antigen. Bars indicate the standard deviation. HBsAg: Hepatitis B Surface Antigen, Arch: Archaeosome, C/IFA: Complete/Incomplete Freund’s Adjuvant
|
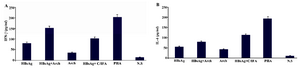
Figure 2. Analysis of IFN-γ and IL-4 level secreted by immunized mice splenocytes. Splenocytes of mice were isolated two weeks after the last injection, stimulated by either HBsAg (10 µM) or PHA (5 µg/ml) for three days and finally IFN-γ; A) or IL-4; B) levels were measured. Bars indicate mean values±SD of three independent experiments. HBsAg: Hepatitis B Surface Antigen, Arch: Archaeosome, C/IFA: Complete/Incomplete Freund’s Adjuvant, PHA: Phytohemagglutinin, N.S: Normal Saline
|

Figure 3. Analysis of IFN-γ and IL-4 secretion frequency by ELISpot assays. Splenocytes (1×105cells/well) from individual immunized mice were cultured in the presence of HBsAg (10 µM) for 40 hr at 37oC in 5% CO2. Spot-forming cells (SFCs) corresponding to the number of IFN-γ and IL-4-secreting splenocytes were counted under a dissection stereoscope. PHA (5 µg/ml) and an irrelevant peptide (aa 132-145 HCV-Core) were applied as a positive and negative control, respectively. Data are shown as means±standard errors per groups and indicate the significant differences (*p<0.05 and **p<0.01) according to Mann-Whitney non-parametric analysis. HBsAg: Hepatitis B Surface Antigen, Arch: Archaeosome, C/IFA: Complete/Incomplete Freund’s Adjuvant
|

Table 1. Formulation of M. smithii specific medium recommended by DSMZ
|
|