New Variations in the Promoter Regions of Human DOCK4 and RAP1A Genes, and Coding Regions of RAP1A in Sporadic Breast Tumors
-
Jalali, Akram
-
Genetic Research Centre, University of Social Welfare and Rehabilitation Sciences , Tehran, Iran
-
Ebrahimi, Hassan
-
Genetic Research Centre, University of Social Welfare and Rehabilitation Sciences , Tehran, Iran
-
Ohadi, Mina
-
Genetic Research Centre, University of Social Welfare and Rehabilitation Sciences , Tehran, Iran
-
Karimloo, Masood
-
Epidemiology and Biostatistics Department, University of Social Welfare and Rehabilitation Sciences , Tehran, Iran
-
Irani Shemirani, Atena
-
Reproductive Biotechnology Research Centre, Avicenna Research Institute, ACECR , Tehran, Iran
-
Mohajer Maghari, Behrokh
-
Reproductive Biotechnology Research Centre, Avicenna Research Institute, ACECR , Tehran, Iran
-
 Khorram Khorshid, Hamid Reza
Ph.D., Genetic Research Centre, University of Social Welfare and Rehabilitation Sciences, Tehran, Iran, Tel: +98 21 22180084-92, E-mail: hrkk1@uswr.ac.ir
Khorram Khorshid, Hamid Reza
Ph.D., Genetic Research Centre, University of Social Welfare and Rehabilitation Sciences, Tehran, Iran, Tel: +98 21 22180084-92, E-mail: hrkk1@uswr.ac.ir
-
Genetic Research Centre, University of Social Welfare and Rehabilitation Sciences , Tehran, Iran
-
Reproductive Biotechnology Research Centre, Avicenna Research Institute, ACECR , Tehran, Iran
Abstract: Breast cancer is the most common cancer among women in developed countries. The prevalence of the disease is increasing in the world. Its annual incidence among Iranian women is about 7000 cases. RAP1A, a tumor suppressor gene, is located at 1p13.3 and plays an important role in the cellular adhesion pathway and is involved in the pathogenesis of breast cancer. The DOCK4 gene, which is located at 7q31.1, specifically activates RAP1A gene. In the present study, DNA samples from 64 cases of sporadic breast tumors (referred to Mehrad Hospital in Tehran) were screened using PCR-SSCP method and the number of observed variations compared with the control group (100 normal women). Mutation detection for coding exons of RAP1A gene and the 500 bp upstream of transcription initiation site as promoters of both DOCK4 and RAP1A were carried out and compared with the control group. The promoter region of DOCK4 showed a heterozygous mutation with G>A transition at nucleotide -303 in a fibroadenoma case. With regard to RAP1A we found a heterozygous mutation, G>A transition in an adenoid cystic carcinoma case, and another heterozygous mutation, G>T transversion in an intraductal papilloma case both at nucleotide +45. A homozygous variation, T>A transversion was also found at nucleotide +29 of a fibroadenoma case. The differences in the frequency of variations mentioned above were not statistically significant. However Fisher’s exact showed significant difference for T>A transversion. Although, the higher frequency of these mutations and variations may be related to the disease, a larger sample size is needed for the confirmation of our findings.
Introduction :
Breast cancer is the most prevalent cancer among women and is the second cause of cancer death in developed countries. In North America and Europe 25% of total cancers in women belong to breast cancer (approximate-ly 20% of women deaths). The prevalence of this disease is increasing to the extent that 1/8 women are possibly affected (1). Among Iranian women, breast cancer incidence rate is about 1/10-1/12 and is the first leading cause of cancer death. Different studies in Iran indicate that the incidence of breast cancer in women is 22 per 100,000. The prevalence in the same population was 120 per 100,000 (2). Breast cancer is classified into two types in terms of original cause of the disease; hereditary and sporadic. Five to ten percent of total breast cancers are hereditary (familial) and the rest are sporadic.
Mutations in BRCA1 and BRCA2 account for about 80% of the hereditary type. Further-more, P53 gene mutations which occur in 50% of human cancers are known to be one of the main causes of breast cancer (3). Tumor Suppressor Genes (TSG) play important roles in different stages of cell cycle and in inhibition of uncontrolled cell proliferation. It has been known that alteration of these genes contribute to sporadic breast cancers.
When both alleles of these genes are inactivated, the cells become cancerous. Loss of heterozygosity (LOH) is one of the phe-nomena that inactivates the alleles of these genes (4).
DOCK4 and RAP1A genes are located at chromosomal loci in which high frequency LOH is observed. The protein expressed by RAP1 gene is a small GTPases which belongs to the RAS super-family and shows 50% amino acid sequence homology with RAS (5).
In addition to the main functions of RAS and RAP1 in cell proliferation and differenti-ation, RAP1 also plays some important roles in the regulation of cell adhesion in epithelial tissue (6).
Cytogenetic abnormalities on the short arm of chromosome 1, i.e. 1p13, are the most prevalent chromosomal aberrations in breast cancer. LOH studies show frequent deletions in 1p22, 1p13, 1p31-32.2 loci in relation to breast cancer. Structural changes in chromo-some 1 especially disarrangements in band 1p13 are the common cause of cytogenetic abnormalities in breast cancers. Mitchel et al showed that 20% of 113 breast tumors contain LOH in 1p13 locus (5). In this context, Bieche et al demonstrated the involvement of two regions in chromosome 1p deletion in breast tumors. One of these deletions harbor Dock4 gene. In his study, using D1S457 and DIS453 markers, the frequency of LOH occurrence was estimated at 34.3% and 21.8%, respect-ively. Considering the fact that the afore-mentioned markers are located at 1p13 locus, these findings most probably indicate that one or two suppressor genes reside at 1p32 band where LOH has been frequently observed in human breast cancer (7, 8).
The DOCK4 gene, a member of the CDM gene family, is located at chromosomal locus 7q31 and is involved in activation and regula-tion of small GTPase protein family including RAP1. Therefore, DOCK4 is a tumor suppres-sor gene and encodes a protein which regula-tes and affects intercellular junctions through activation of RAP1 and is disrupted during tumorigenesis (9).
Marker D7S523 on chromosome 7q31 is frequently deleted due to LOH, in human prostate, ovarian and breast cancers (10). This marker is located at the first intron of DOCK4 (11). Schmutzler et al and Sztanm et al showed that a high frequency of allelic loss (LOH) at D7S523 marker in human breast cancer indicates the presence of a tumor suppressor gene at this region (12, 13). In another study by Balkhi et al, it was revealed that DOCK4 gene was not expressed in 53% of patients with breast cancer. The difference between patient and control groups was significant (14). This result indicated that DOCK4 gene expression is disrupted in breast cancer patients. The promoters are involved in gene expression regulation, warran
Materials and Methods :
This research is a series of case studies. The groups studied (64 cases) were randomly selected from women having breast tumors who were admitted in Mehrdad hospital from October 2004 to December 2005. The criter-ion for inclusion was the pathological con-firmation of breast cancer diagnosis (sporadic invasive ductal breast cancer) or the presence of benign breast tumor. The criterion for exclusion consisted of the existence of breast cancer in the family. For all patients a ques-tionnaire regarding their clinical and patho-logical history was completed. The results in case group were compared with 100 normal and healthy women (control group).
Consent forms were signed by all patients. All samples were analyzed while they were unknown to the examiner. All related infor-mation about DOCK4 gene such as complete gene sequence, presence of SNPs in exons, mRNA and protein sequences were collected from Ensembl and NCBI related data bases (15, 16).
Around 500 bp upstream of transcription initiating site was considered as promoter region and analyzed for the presence of pro-moter-specific sequences using McPromoter promoter prediction program (17). Multiple alignment of human and mouse (Mus mus-culous) Dock4 genes were carried out using DiAlign multiple alignment program (18). Transcription factors binding sites in both mouse and human promoter regions were analyzed and compared by ConSite program (19). Tandem repeats were analyzed using Tan-dom Repeats Finder Program (20). Using Pri-mer3 program, 3 sets of overlapping primers were designed for 500 bp upstream of trans-cription start site as promoter region of DOCK4 and RAP1A gene (21).
For each coding exon of RAP1A gene a pair of primers extending from the flanking introns was designed as well. DNA was ex-tracted from tumoral tissues by both standard method (phenol/chloroform) and DNA extrac-tion kit (DNPTM, Cina Gene) according to manufacturer's instructions. DNA from blood was extracted using salting out method. Primer sequences are shown in tables 1 and 2.
PCR reaction was performed in a total volume of 25 µl in 0.2 ml Eppendorf tube using Eppendorf thermal cycler. The reaction mixture consisted of 30 ng DNA template, 10 pmol of each primer set (constructed by TAG Copenhagen), 0.3 U of Taq polymerase (Supper Taq), 1.5 Mm MgCl2 and 0.2 Mm dNTPs (Merck). The denaturing, annealing and the extension-time length as well as the temperature for each set of primers were opti-mized and are shown in table 3. Initial dena-turation and final extension time and tempera-ture were (3', 94 ?C) and (7', 72 ?C), respect-ively.
First, PCR products were run on 8% acryl-amide gel to observe the desired band at the right size. To perform SSCP analysis, PCR product obtained from DOCK4 -Pro1& 2 and also RAP1-Ex 3& 4 were run on 10.5% acryl-amide gel and other PCR products were run on 9.5%. Once acryl-amide gel was made, it was kept in the fridge for 1 hr to reach 4 ?C temperature. Then, gel was put on SSCP tank. PCR products were mixed with an equal volume of loading buffer (PCR gel electro-phoresis loading buffer and 90% formamide). Tubes were immediately put on mixture of crushed ice and water for 5 min and after-wards the prepared samples were run on SSCP gel. Marker with the quantity of 150 ng was loaded along side the gel. Table 4 shows the running condition of SSCP gel.
To determine any nucleotide change, the shifted samples on SSCP gel along with normal sample from the same PCR product were sequenced using Chain Termination Method (Microgene Co).
Result :
Bioinformatical analysis of the DOCK4 gene
DOCK4 contains 53 exons and encodes a protein consisting of 1954 amino acids. DOCK4 has 5 domains and is an intramem-brane protein. The Dock4 protein is a member of Dock protein family (DOCK1 to DOCK11) which all contain dedicator of cytokinesis motif. This motif consists of 200 amino acids and has guanyl-nucleotid exchange factor activity. Frequent splicing results in 6 mRNAs coding for 6 protein isoforms (15, 16). This gene contains 9 SNPs in exons 5, 9, 17, 24, 44 and 52 which results in amino acid changes in constructed proteins (15). No pro-moter-specific sequence was recognized in 500 bp upstream of the transcription initiation site using McPromoter promoter prediction program (17). Mouse and human sequences alignment of this region using DiAlign pro-gram showed 88% homology (19). No homo-logy was observed between transcription factors binding sites in mouse and human Dock4 gene using Consite program (19). Also, Tandem repeats finder program could not find any tandem repeat at either promoter or at 5' upstream or 3' downstream regions of the gene (20).
Mutation analysis of the genes PCR products were run on %8 acrylamide gel and were observed after silver nitrate staining (Figure 1).
SSCP analysis of the samples
There was no band shift in sequences obtained from primer sets DOCK4, PRO2 and PRO1. However, one band shift in DOCK4, PRO3 sequence was found. The sample was from a patient who had fibroadenoma breast mass. Sequencing analysis showed a C>G heterozygote variation. This variation is lo-cated at position -303 (303 nucleotide up-stream of the transcription start site) and +224 of the PCR product sequence (Figure 2). No shift was observed in control group samples.
No band shift was observed in PCR products obtained from RAP1A exon (3, 4 and 6) and promoter regions. Three shifts were observed in RAP1exon3 sequence which all belonged to introns. Sequencing analysis of PCR product from sample 4 (obtained from a patient with cystic adenoid carcinoma) showed a heterozygote mutation with G>A transition. This variation was located at position +45 from the intron starting point and +146 from 5' site of the PCR product. Sequence analysis of sample 8 that belonged to a patient with breast tumor (intraductal papilloma) showed a heterozygote mutation (G>T transversion) at the same site as it was observed in sample 4. T>A transversion was detected from sample 6 obtained from a patient with fibroadenoma. This variation is located at position +29 from the starting site of the intron and +130 from the 5' region of the PCR product (Figure 3). SSCP analysis of the control samples however did not reveal any band shifts.
Statistical analysis
Fisher's exact test showed that C>G variation in DOCK4 gene is independent from the type of the disease (p=0.13). Proportion of C>G mutation was higher in patients with breast tumor than the normal group. However this difference was not statistically significant (RR= 14.22, 95% CI = 0.7-842).
Fisher's exact test showed that occurrences of RAP1A gene with G>T and T>A variations (p= 0.13) as well as G>A mutation (p=0.329) are independent from the type of the disease. Although the proportion of patients carrying G>A mutation was also higher in cases with breast tumor compared with the normal group, the increase was not statistically significant (RR=4.17, 95% CI = 0.21-247).
Furthermore, we found one case of homozygote variation (two allelic variations, T>A) among 15 cases of benign breast tumor. This is equivalent to two variations among 30 alleles. On the other hand no variation of this type was found among 100 normal subjects in the study which is equivalent to 200 alleles. When we compared the two groups using the Fisher exact test, the observed difference was statistically significant (p=0.017).
Discussion :
In addition to important roles of RAS and RAP1 in cell proliferation and differentiation, RAP1 regulates cell adhesion in epithelial tissues (6). LOH studies on chromosome 1 in different malignancies exhibit frequent deletions in 1p22, 1p13 and 1p31-32.2 loci. Structural chromosome1 changes, specifically disarrangements in band 1p13 are the common cause of cytogenetic abnormalities in breast cancers (5).
DOCK4 gene, a member of CDM gene family, resides at long arm of chromosome 7 (7q31) and is known to be a tumor suppressor gene. DOCK4 is involved in activation and regulation of small GTPase protein family specifically RAP1, and hence stimulates the formation of intercellular junctions. DOCK4 mutations found in prostate and ovarian cancers prevent RAP1 GTPase activation. In this regard, reduction of DOCK4 expression in breast cancer patients has been reported (14). Considering the function of DOCK4 and RAP1 genes and also their location at a locus of a chromosome in which high frequency LOH has been reported for the majority of breast cancers, it can be deduced that these two genes are tumor suppressor genes.
In this study promoter analysis of DOCK4 gene in a benign tumor sample (fibro-adenoma) revealed a heterozygote mutation C>G at position -303 (6.66%). Gene analysis showed three variations in the third intron of RAP1 gene as follows: heterozygote mutation G>A (2.04%) in an adenoid cystic carcinoma at position +45, heterozygote variation G>T (6.66%) in a benign tumor (intraductal papil-loma) at position +45 and a homozygote variation T>A (6.66%) in a benign tumor (fibroadenoma) at position +29. Statistical tests showed that variations and mutations in both genes are independent from the type of the disease. However, considering the allelic frequency of T>A this variation was observed just in 15 cases of benign breast tumor which is equivalent to two variations among 30 alleles. On the other hand no variation of this type was found among 100 normal subjects in the study which is equivalent to 200 alleles. Therefore its occurrence possibly is related to the type of the disease. Since no heterozygote variation of this type was found in case and control groups it should be considered that probably SSCP method is not able to detect the heterozygote variant and other screening methods like RFLP, ASO or sequencing should be applied to determine the exact fre-quency of this variation.
In summary DNA samples from 64 cases of sporadic breast tumors were screened and the observed variations in DOCK4 and RAP1A genes were compared with the control group (100 normal women). Although this study did not reveal any decisive pathological mutation but it uncovered four variants at low fre-quency that one of them was new and unclas-sified (G>A variant in a benign tumor sample at position +45 of the third intron of RAP1A gene). As a result, performing new studies to decipher the significance of these variants is indicated.
Acknowledgement :
We would like to express our sincere grati-tude to all principal investigators and research assistants of Genetic Research Centre, Social Welfare and Rehabilitation Sciences Univer-sity, who have contributed to this study.
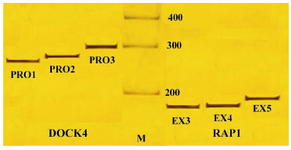
Figure 1. PCR amplified products of DOCK4 and RAP1A genes, M: 100 bp DNA ladder; PRO1, PRO2 and PRO3: Different regions of DOCK4 gene promoter; EX3, EX4 and EX5: PCR amplified fragments (exons 1, 2 and 3) of RAP1
|

Figure 2. DOCK4 and PRO3 PCR products of patient's samples, M: 100 bp DNA Ladder; S1-S7: samples; The PCR product size is 284 bp and single strand is separated in 500 bp and 560 bp. In sample 5, an obvious band shift is observed
|

Figure 3. RAP1A. EX3 PCR products from patient's samples, M: 100 bp DNA Ladder marker; S1-S11: RAP1.EX3 PCR products from different samples; the size of PCR products are 174 bp. Two band shifts in single-strand sequences are observed at positions around 290 and 340 bp. There are 2 faint bands of single-strand sequences at positions around 265 bp and 315 bp which are not found in S4 and S8. All bands are more intense in S6
|
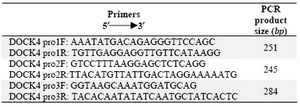
Table 1. Primer sequences for the amplification of DOCK4 promoter region
|

Table 2. Primer sequences used for the amplification of promoter and exon-coding regions of RAP1A gene
|
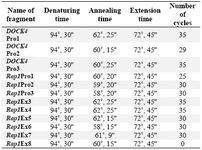
Table 3. The amplification condition
|
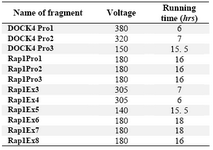
Table 4. Voltage and time of electrophoresis on SSCP gel for each PCR product
|
|