A Novel Approach for High Level Expression of Soluble Recombinant Human Parathyroid Hormone (rhPTH 1-34) in Escherichia coli
-
Hamedifar, Haleh
-
Research and Development Department, CinnaGen, Simindasht Industrial Area, Karaj, Iran
-
Salamat, Firoozeh
-
Research and Development Department, CinnaGen, Simindasht Industrial Area, Karaj, Iran
-
Saffarion, Mohammad
-
Research and Development Department, CinnaGen, Simindasht Industrial Area, Karaj, Iran
-
Ghiasi, Mohammad
-
Research and Development Department, CinnaGen, Simindasht Industrial Area, Karaj, Iran
-
Hosseini, Alireza
-
Research and Development Department, CinnaGen, Simindasht Industrial Area, Karaj, Iran
-
Lahiji, Hadi
-
Research and Development Department, CinnaGen, Simindasht Industrial Area, Karaj, Iran
-
Nouri, Zomorrod
-
Research and Development Department, CinnaGen, Simindasht Industrial Area, Karaj, Iran
-
Arfae, Hamed
-
Research and Development Department, CinnaGen, Simindasht Industrial Area, Karaj, Iran
-
 Mahboudi, Fereidoun
Research and Development Department, CinnaGen, Simindasht Industrial Area, Iran, Tel: +98 21 42915000; Email: mahboudif@cinnagen.com
Mahboudi, Fereidoun
Research and Development Department, CinnaGen, Simindasht Industrial Area, Iran, Tel: +98 21 42915000; Email: mahboudif@cinnagen.com
-
Research and Development Department, CinnaGen, Simindasht Industrial Area, Karaj, Iran
Abstract: Background: Parathyroid hormone (PTH) secreted by parathyroid glands regulates the metabolism of calcium and phosphorus in bone and kidney. Thereby, it can stimulate bone formation, and is a promising agent in the treatment of osteoporosis. Mature form of PTH consists of 84 amino acids; however, the first 34 residues of PTH cover the majority of hormonal action.
Methods: In this study, the fusion form of highly soluble rhPTH was expressed at high level in Escherichia coli (E. coli). His6-thioredoxin as an extension for rhPTH improves the solubility of inclusion body. His6-thioredoxin-hPTH (1-34) was ligated into pET32a expression vector. The insertion of 5 amino acids (Asp-Asp-Asp-Asp-Lys) in the N-terminal of PTH made this protein to be digestable specifically by enterokinase enzyme. The fusion form of rhPTH was harvested and purified by immobilized affinity chromatography followed by digestion with enterokinase. Digested rhPTH was purified by applying on size exclusion and ion exchange chromatography to get the highest purity.
Results: The mass spectroscopy analysis shows rhPTH molecular weight was 4117.5 Da. The purity was measured by HPLC column which showed more than 97%. Bioassay analysis of rhPTH was performed on rat sarcoma cell UMR-106 in parallel with commercially available rhPTH, Forteo. The result was measured through immunofluorescence detection kit. The data showed that the potency of rhPTH was comparable with commercially available medicine.
Conclusion: Thioredoxin was applied as a fusion partner for production of highly soluble rhPTH. This specific fusion partner increased protein solubility and decreased protease reactivity. Purification process was optimized for high recovery and for purity more than 99%. As its biological activity is comparable with marketed drug, this protein is qualified for biopharmaceutical usage.
Introduction :
Extracellular calcium homeostasis is regulated by Parathyroid hormone (PTH), which acts directly on kidney and bone. In human, PTH is an 84-amino acid polypeptide 1. The intact protein is composed of 115 amino acid residues. Twenty five N-terminal amino acids are cleaved off when they pass through the lumen of endoplasmic reticulum by signal peptidase 1,2. Other 6 amino acid residues are cleaved off in Golgi complex and mature PTH with 84 amino acids is secreted 2. It is shown that the first 34 N-terminal amino acids cover the most hormonal action of the intact hPTH. This fragment can regulate calcium and phosphate homeostasis and control bone resoprtion as well 3. Daily subcutaneous injection of human PTH (hPTH) (1-34) stimulates bone formation and intensifies bone mass in osteoporosis 4 and ovariectomized monkeys 5. Moreover, it is demonstrated that hPTH (1-34) possess anti-apoptotic effects in rat osteosarcoma 6. These findings reveal that hPTH (1-34) has medical and pharmaceutical applications for osteoporosis treatment 7. These may be due to the fact that hPTH stimulates cyclic AMP-dependent protein kinase in a group of bone cells bearing PTH receptors and indirect osteoblasts proliferation 8.
hPTH(1-34) has been produced in different forms including synthetic and recombinant. Variety of host systems including E. coli, Saccharomyces cerevisiae, Pichia pastoris, and mammalian cell lines have been used to express this protein 9,10. Comparing the full length hPTH, recombinant N-terminal fragment (amino acids 1-34) [rhPTH (1-34)] has been shown to have most of the hormonal actions in body 10.
One of the obstacles in expression of recombinant hPTH (rhPTH) is digestion by intracellular proteases 11. A successful scenario to overcome this issue is to produce rhPTH in the fusion form with another polypeptide. There are several reports for application of different fusion partners including Achromobacter protease I, Staphylococcus aureus V8 protease 12 and Fc fragment of human IgG113. b-galactosidase derivatives are examples of other reported fusion partners 12. In addition, Chunxiao and his/her co-worker selected the N-terminal Pro-Pro as the fusion partner 14,15.
In this article, thioredoxin was selected as the extension for rhPTH. Thioredoxin makes the expressed protein more soluble and the inclusion body is dissolved in lower urea concentration 16. In addition, it is possible to obtain higher expression by using this fusion partner and it can be purified using IMAC regarding His6 tag binding affinity to N terminal of the expression cassette 16. Liu and co-workers expressed PTH (1-84) using the same fusion partner in E. coli 10. However, there is no report on recombinant hPTH (1-34) expression in E. coli with present fusion partner design, including specific protease recognition site (Asp-Asp-Asp-Asp-Lys). This fusion polypeptide prevents non-specific digestion of the protein and enables an efficient purification process.
The purification optimization was performed by using three steps with Ni-NTA resin, size exclusion and ion exchange chromatography using SP sepharose resins. The purification results were analyzed by SDS-PAGE electrophoresis and confirmed by reverse phase analytical high performance liquid chromatography (RP-HPLC).
Materials and Methods :
Strains, plasmids and enzymes:E. coli BL21(DE3) (Novagen, USA) was used as the host for gene expression. The pET32a(+) expression plasmid was obtained from Novagen, USA. Restriction enzymes and modifying enzymes were purchased from Takara, Japan and enterokinase was obtained from Invitrogen, USA.
Construction of the expression vector:The gene encoding the first 34 amino acids of PTH, 5 amino acid residues (Asp-Asp-Asp-Asp-Lys), thioredoxin and His6 tag sequence, were synthesized by CinnaGen, Iran. PTH(1-34) gene was located at the C terminal of Lys. This sequence was ligated into pET32a+ (Invitrogen Co, USA) between Bg1 II -Kpn I site of plasmid, then the pET32a-PTH was transformed into E. coli BL21 as cited in the instruction manual of the kit (Invitrogen Co, USA) (Figure 1). The orientation of the gene was confirmed by restriction enzyme digestion and DNA sequencing (SinaClone, Iran) (data not shown).
Expression in E. coli:A single colony of the fresh E. coli BL21 transformants bearing pET32a-PTH was cultured in 10 ml LB media containing Ampicilin (100 µg/ml) overnight 17. The cell suspension, 1 ml, was transferred to 300 ml of the same media for an additional 18 hr at 37C and 160 rpm. The harvested cell suspension was diluted 10-fold in fermentation process with LB Broth containing Ampicilin (100 µg/ml) and cultured in 5L fermentor at 37C with aeration rate of 1 (v/v/m) and agitation rate of 200-600 rpm. The expression of the His6-Thioredoxin-hPTH (1-34) fusion protein was induced at OD 0.9 by addition of IPTG (CinnaGen Co., Iran) to a final concentration of 1 mM. The biomass was harvested after 5 hr of fermentation followed by centrifugation at 9500 rpm for 10 min. The cell pellet was kept frozen at -70C. Polyacrylamide-gel electrophoresis in the presence of SDS (SDS-PAGE) was used to determine the expression level of the His6-thioredoxin-hPTH (1-34) fusion protein.
Extraction and site-specific cleavage of product:The frozen biomass of E. coli was resuspended in lysis buffer (20 mM Tris-HCl pH=8 containing 6 M Urea) on ice. The suspended cells were disrupted and homogenized at 1200 Psi, followed by centrifuging at 9500 rpm, 4C for 45 min. The supernatant containing the fusion protein was collected and applied to Ni-NTA resin (column XK 50/40, GE, USA). The column was equilibrated with 50 mM Tris-HCl pH=8 containing 6 M Urea followed by washing with wash buffer (20 mM Tris-HCl pH=8 containing 6 M Urea, 2 M NaCl). The fusion protein was eluted with elution buffer (20 mM Tris-HCl pH=8 containing 6 M Urea, 2 M NaCl and 18 mM Immidazole). Fractions containing His 6-Thioredoxin – hPTH (1-34) fusion protein were pooled and desalted by dialyzing for 24 hr at 4C in dialysis buffer (20 mM Tris-HCl pH=8 containing 10% w/w Tween 20 ,0.1 M CaCl2).
Proteolytic cleavage of fusion protein with enterokinase:In order to cleave hPTH from its tag, the desalted fusion protein was digested with enterokinase enzyme (EK MAX, Invitrogen Co., USA) for 16 hr at room temperature with different enzyme units. The digestion reaction was terminated by reducing the pH with 2 M acetic acid.
Purification of recombinant PTH:The digested mixture was applied to size exclusion chromatography column (GE, USA) by FPLC (AKTA purifier, GE, USA) that was equilibrated with 20 mM sodium acetate and 50 mM NaCl Fractions were analyzed by SDS-PAGE. Then hPTH without His6-thioredoxin tag was pooled and loaded on ion exchange chromatography column with SP sepharose resin (GE, USA). The column was equilibrated and washed with equilibration buffer (0.01 M ammonium acetate pH=4.5 containing 50 mM NaCl) and washing buffer (0.02 M ammonium acetate pH=5.2 containing 0.1 M NaCl), respectively. The recombinant hPTH (1-34) peptide was eluted with 0.02 M Ammonium Acetate and 1 M NaCl. The fractions were collected and kept at 4C.
Purity analysis:To estimate the purity of the SP fractions, RP-HPLC analysis (C18, Agilant, USA) was performed. HPLC chromatogram of hPTH (1-34) was performed using a linear gradient of 100% A, 0% B solutions in 25 min, flow rate 1 ml/min. Solution A contained acetonitril and 0.2 M Na2SO4 (1:9 w/w) and solution B contained acetonitril and 0.2 M Na2SO4 (1:1 w/w). Sample, rhPTH (1-34), and standard hPTH (Forteo) (Eli Lilly, France) were diluted with dilution buffer (acetonitril and 0.3 M Na2SO4, 31/69 w/w) to the final concentration of 0.1 mg/ml. Then 25 µl of each diluted samples were applied to HPLC column. Proteins were monitored by UV detection at 214 nm. Mass spectrometry was performed to obtain the molecular weight of the protein on a Finnigan LCQ-Classic MALDITOF mass spectrometer.
Biological activity:Preparing sample dilution: Samples containing Forteo (Eli Lilly, France) and rhPTH were diluted to final concentration of 1 µM/ml with DMEM-F12,1% BSA(Bovine Serum Albumin) (Sigma, USA). Then, a series of dilutions, 1:1000-1:8000 final concentration in the assay, were prepared with DMEM with BSA 1%.
Standard preparation:The 30 µM cAMP calibrator (Sigma, USA) was diluted with cAMP Assay Buffer (Catch Point Cyclic-AMP Fluorescent Assay Kit, Molecular Devices, USA) to prepare stock calibrators of 1:1000-1:8000 final concentration in the assay.
Cell preparation:Flasks containing UMR 106 cells (ATCC,USA) were treated with 1 ml trypsin (GIBCO, USA) and resuspended to final concentration of 0.1×106 cell/well with DMEM-F12 (GIBCO,USA), 10% FBS (GIBCO, USA). Then 200 µL of cell solution, 20000 cell/well, were transferred to each well of 96 well-plate and incubated for 24 hr. Wells were washed two times with HBSS buffer (Hank’s Balanced Salt Solution). Afterwards, 100 µl of each hPTH (1-34) and Forteo, reference standard, (Eli Lilly, France) dilutions were separately added to each well and incubated with gentle shaking at 25°C for 20 min. Samples were discarded and wells were washed twice with 300 µl HBSS. Assay Lysis Solution, 100 µl (Catch Point Cyclic-AMP Fluorescent Assay Kit, Molecular Devices, USA) were added to each well and plate was incubated for 30 min at 37°C.
Cell stimulation and cAMP detection:Cell lysate was mixed gently with multichannel pipette and 40 µl of the solution was transferred to cAMP screen assay plate. Reconstituted Rabbit anti-cAMP antibody, 40 µl, was added to each well except control wells. The plate was placed on shaker for 5 min. Then 40 µl of reconstituted HRP-cAMP conjugate was added to all wells and incubated for 2 hr at room temperature. The plate content was aspirated and washed 4 times with 300 µl wash buffer (Catch Point Cyclic-AMP Fluorescent Assay Kit, Molecular Devices, USA) for each wash. Then 100 µl of Spotlight Red substrate (Catch Point Cyclic-AMP Fluorescent Assay Kit, Molecular Devices, USA) were added to each well. The plate was covered to be protected from light and was incubated in room temperature for 10 min. Fluorescent intensity of the plate was read in Excitation filter 525-530 nm, Emission filter 590-570 nm. The average intensity values (y-axis) were plotted against calibrator concentration (x-axis) to create a calibration or standard curve. To calculate cAMP doses, hPTH samples were interpolated using this curve.
Results :
Expression construct for recombinant hPTH:As shown in figure 1, the gene sequence encoded for amino acids 1-34 from PTH was inserted in the C terminal portion of thioredoxin gene. The DNA sequence for 5 amino acids (Asp-Asp-Asp-Asp-Lys) as a specific recognition site was inserted between PTH gene sequence and the fusion partner DNA sequences. His-tag was added to the downstream of Bgl II site. Two restriction enzyme recognition sequences, BglII and Kpn I, were added to the N terminal and C terminal of the fusion protein, respectively. The construct was inserted into pET32a(+) resulting in pET32a(+)-hPTH34 plasmid.
The expression vector subsequently was transformed into E. coli BL21 (DE3) to form the engineered bacterium. The highest level of expression was obtained 5 hr after induction with IPTG. This level of expression usually makes the recombinant protein hard to solubilise and form inclusion bodies. His6-Trx fusion partner has beneficial features of high solubility and it provides the possibility of using affinity chromatography for purification. As shown in figure 2 (lane 1), there is low expression in pre-induction sample. However, the productivity was higher three hours after the induction and reached to the highest level after 5 hr (Figure 2, lane 7).
Cleavage of the product:The fusion protein was purified using Ni-NTA due to the presence of His6 tag. Then, the fusion partner with 18 kDa, was separated from rhPTH through digestion with Enterokinase (Figure 3). The enzyme to substrate ratio was optimized at lowest level, 0.341 per/mg of protein. Two bands, fusion partner 18 kDa and rhPTH 4 kDa, are clearly demonstrated in the SDS-PAGE. Lane 1 is the undigested fusion protein. Lane 2 is rhPTH standard. Lane 3 shows complete digestion fusion protein. Presence of specific digestion site (Asp-Asp-Asp-Asp-Lys) for enterokinase prevents non-specific digestion of rhPTH.
Purification process:The digested proteins were applied to size exclusion column (Superdex 75) followed by SP sepharose column. The purified rhPTH with more than 97% purity was obtained (Figure 4A: lanes 2, 3, 4, 5, 6, 7 and 8 different fractions from ion exchange column). These findings are in line with what other researchers have previously found 12. In our system the purification steps were restricted to three columns. The majority of the impurities were taken during Ni-NTA followed by specific binding to Sp sepharose fast flow column. The final recovery found to be 42% (data not shown) with 99.7% purity. The first 8 N-terminal amino acids of the purified proteins were similar to what was found in standard rhPTH from NIBSC through mass spectroscopy (data not shown).
The purity of rhPTH was confirmed using RP analysis by analytical HPLC (Figure 5). A single sharp peak was demonstrated at retention time of around 20 min. The RP HPLC diagram was the same as the standard sample from NIBSC (data not shown) and the impurities were less than 0.3%. In addition, Mass spectrophotometry analysis show that the molecular weight of rhPTH was 4117.5 Da (Figure 6). The molecular weight of pro-pro-pro PTH was reported as 4311.46 14. If Pro-Pro-Pro is eliminated, the right molecular weight (4117.5 Da) will be observed.
Biological activity:The binding of the rhPTH receptors was measured by assaying stimulated cAMP generation by the osteogenic sarcoma cell culture. PTH bioassays in vitro have the most commonly utilized cultured animal tissue, and the tissue extract contains adenlate cyclase and animal cells 18. PTH binds to its receptors causing a complex response in tissues, cells or extracts that can be measured by the stimulated generation of cAMP 19,20. In the present study, UMR 106 cells were chosen for in vitro determination of rhPTH biological activity.
The purified rPTH increased the cAMP level in UMR 106 cells which is rat osteogenic sarcoma cell line by interaction with surface receptors (Table 1). The positive control was the commercially available rPTH, Forteo. The potency of the sample was 99.7% according to the standards and the testing criteria for protein activity. As it is shown in table 1, the optical density obtained in each dilution was comparable with what was obtained by Forteo. The standard curve demonstrates validation of the test. The potency of rhPTH was similar to Forteo.
Discussion :
The successful expression of rhPTH in E. coli needs fusion partner, since low molecular weight rhPTH without fusion partner is susceptible to endogenous protease digestion 11. In addition, the problems encountered in over expression of other proteins in bacteria are overcome through the use of fusion proteins 21. Therefore, selection of the right fusion partner is a critical issue. Recombinant hPTH (1-34) has been expressed in E. coli by fusing to different fusion partners such as asparginase C-terminal fragment and human growth factors 22, the E. coli maltose-binding protein (MBP, 40 kDa), the E. coli N-utilizing substance A (NusA, 54.8 kDa), and thioredoxin (Trx, 11.7 kDa) 10.
E. coli Thioridoxin (Trx) is an abundant small (Mr 12,000) redox protein and a catalyst for reducing protein disulfide bonds 23. Trx takes less energy to obtain a certain number of molecules and reduce steric hindrance, therefore it reduces inclusion body and simplifies downstream applications 24. It also has been reported that Trx increases the solubility of foreign proteins expressed in E. coli as well 24.
Liu and colleagues showed that the expression level of 23% of total bacterial protein is high enough for large scale production 10. In the present study, the level of expression was more than 50% of total bacterial protein.
Since rhPTH contains Lys, Arg, Glu, Asp and some aromatic amino acids, the selection of a protease that recognizes these amino acids should be avoided. One of the protease specific sites is SVKKR that was ligated in the 5’ site of rhPTH 12. Kex2-660 and kex2 enzymes have been used for the above protease recognition site which contains RR, KR, and RR sequences 8. The recognition sequence, Asp-Asp-Asp-Asp-Lys, integrated in the present construct of rhPTH, is shown to be very specific.
The final yield of rhPTH, after purification, was 100 mg/l, higher than 27 mg/l of the most efficient direct expression strategy 25 and 5-8 mg/l of soluble fusion protein system using the C-terminus of a 15 kDa IgG-binding protein from Staphylococcus aureus protein A as a fusion partner 26, and also more than 30 mg/l of hPTH tagged with GST and cleaved with Factor Xa 27.
Conclusion :
High expression level with high solubility was obtained using thioredoxin as a fusion partner for PTH. This specific fusion partner increased protein solubility and decreased protease reactivity. Purification process was optimized for high recovery and for purity more than 99%. The biological activity and potency were found to be similar to reference standards, Forteo and NIBSC rhPTH. This protein has all the criteria as a good candidate for biopharmaceutical interest since it demonstrates biosimilarity to Forteo, one of the well defined biopharmaceutical proteins which is available in the market.
Acknowledgement :
This study was financially supported by CinnaGen Company. The authors wish to thank Dr. Aghajani for critical reviews and final revisions.
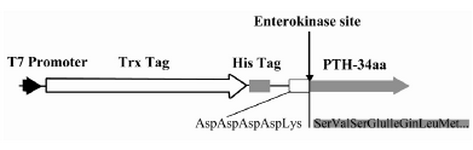
Figure 1. The schematic representation of PTH expression cassette including T7 promoter, thioredoxin coding sequence, His Tag coding sequence, enterokinase recognition site and PTH start coding sequence
|
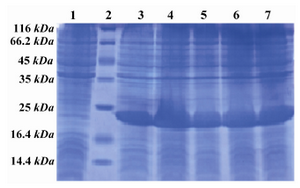
Figure 2. SDS PAGE pattern of induction and expression rate of hPTH (1-34) fusion protein. Lane 1: Before induction, Lane 2: Low MW marker, Lane 3: 1 hr after induction, Lane 4: 2 hr after induction, Lane 5: 3 hr after induction, Lane 6: 4 hr after induction, Lane 7: 5 hr after induction
|
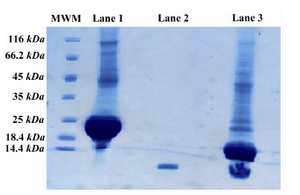
Figure 3. SDS-PAGE pattern of the digested fusion protein. Lane 1 is undigested PTH fusion protein, lane 2 is Forteo as the standard and lane 3 is the digested PTH fusion protein with enterokinase. There are two major bonds after digestion, 18 kDa and 4 kDa represent fusion partner and PTH, respectively
|
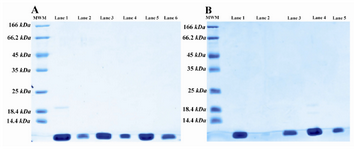
Figure 4. SDS-PAGE pattern of size exclusion (a) and ion exchange (b) chromatography fractions; A) SDS-PAGE analysis of size exclusion column fractions. Lane 1 is the digested sample and lanes 2-6 are fractions of SE column. There is some undigested protein in lane 1; B) SDS-PAGE analysis of SP sepharose FF fractions. Lane 1 is pooled SE column fractions, lane 2 is empty and lanes 3-5 are fractions of SP sepharose FF
|
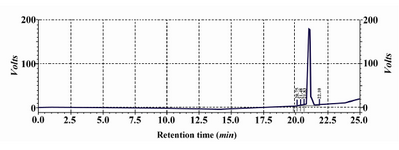
Figure 5. HPLC analysis of SP sepharose pooled fraction. A single peak at 22 min retention time represents the purity of rhPTH (more than 99%)
|
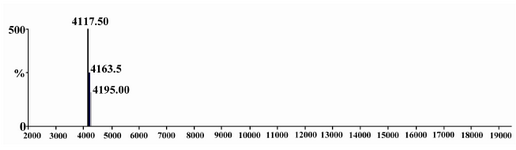
Figure 6. Mass spectrometry of rhPTH done in Boku University, Austria, is showing 4117.50 Dalton molecular weight
|
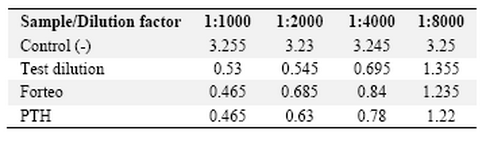
Table 1. Biological activity assay with cAMP production measurement
|
|