Production of Cloned Mice by Nuclear Transfer of Cumulus Cells
-
Kurd, Soleiman
-
Department of Biological Sciences and Biotechnology, School of Science, University of Kurdistan, Kurdistan, Iran
-
Zarei, Mohammad Ali
-
Department of Biological Sciences and Biotechnology, School of Science, University of Kurdistan, Kurdistan, Iran
-
 Fathi, Fardin
Cellular and Molecular Research Center, Kurdistan University of Medical Sciences, Sanandaj, Iran, Tel: +98 871 6131382; Email: farfath@gmail.com
Fathi, Fardin
Cellular and Molecular Research Center, Kurdistan University of Medical Sciences, Sanandaj, Iran, Tel: +98 871 6131382; Email: farfath@gmail.com
-
Cellular and Molecular Research Center, Kurdistan University of Medical Sciences, Sanandaj, Iran
-
Ghadimi, Tayyeb
-
Department of Surgery, School of Medicine, Kurdistan University of Medical Sciences, Sanandaj, Iran
-
Jalili, Ali
-
Cellular and Molecular Research Center, Kurdistan University of Medical Sciences, Sanandaj, Iran
Abstract: Background: Over the past several years, mammals have been successfully cloned by either the splitting of an early stage embryo or nuclear transfer of adult somatic cells (NT) into oocytes. Although it has been 15 years since the generation of the first cloned mammals from somatic cells by NT, the success rate for producing live offspring by this technique is low regardless of the cell type and animal species used. However, these techniques have the potential to be important tools for future research in basic biology. In the present study, we described our experiences in producing successfully cloned mouse using NT method and piezo-actuated micromanipulator.
Methods: B6D2F1 mice, 8-12 weeks old, were superovulated with injections of 5 IU of pregnant mare serum gonadotropin and 5 IU of human chorionic gonadotropin administered 48 hr apart. Enucleation and donor nuclei cumulus cell injection were performed with a piezo-actuated micromanipulator after which activation and trichostatin A treatment were used for reconstructed oocytes. Two-cell stage cloned embryos that developed in the mWM medium were transferred into the oviducts of pseudopregnant NMRI mice.
Results: Of 367 oocytes collected, 131 (69%) developed into 2-cell stage embryos. Of these, 5 (1%) live pups were successfully delivered. We used NMRI foster mother to raise the pups by lactation. One adult cloned mouse was mated, after which she delivered and raised normal offspring.
Conclusion: For mouse cloning, the present study also successfully tested the capability of somatic cell nuclear transfer SCNT using a piezo unit.
Introduction :
Reproductive cloning is defined as the production of a group of individuals with the same genotype by asexual reproduction, a natural occurrence in numerous plants and animals 1. Over the past several years mammals have been successfully cloned by the splitting of an early embryo or Nuclear Transfer (NT) of adult somatic cells into
oocytes. It is difficult to pinpoint the origin of cloning as this technology has been used by numerous scientists. In 1894, Dreisch successfully cloned a sea urchin and several years later Hans Speman successfully cloned a salamander. Briggs and King used the NT technique to clone frogs. In 1985, Steward isolated cells from the roots of a mature carrot, which resulted in the creation of a full clone. Steward also proved that it was possible to generate clones by using somatic cell NT 2. Cloning experiments have also been undertaken by other scientists such as Gurdon (Xenopus laevis) 3 and Willadsen (sheep) 4.
Nuclear transfer techniques for the cloning of domestic animals have undergone rapid development. On July 5, 1996,¬ Wilmut et al used Roslin mehod to clone the first mammal, a sheep named Dolly. In their method, somatic cells were permitted to grow and divide. They were deprived of nutrients and induced into a suspended or dormant stage. An oocyte with removed nucleus was placed in adjacency to a somatic cell and then both cells were shocked by an electrical pulse. Both cells were fused and the oocyte was permitted to grow into an embryo. The embryo is then transferred into a surrogate mother. The birth of Dolly was considered a turning point in the history of cloning in that it showed the feasibility of producing live offspring by somatic cell NT 5. In 1998, Wakayama et al used Honolulu method to inject enucleated oocytes with cumulus cell nuclei and produced the first cloned mouse, Cumulina, which survived 6. They removed the nucleus from a somatic cell and injected it into an oocyte with removed nucleus. The oocyte was bathed in a chemical solution and cultured. The growing embryo was then transferred into a surrogate mother and permitted to develop. Following this success, other animals such as cattle 7, goats 8, dogs 9, domestic cats 10, rabbits 11, horses 12, rats 13, ferrets 14 and zebra fish 15 have been successfully cloned.
In Iran, cloning experiments were performed by Kazemi Ashtiani and colleagues at Royan Institute. They produced the first cloned sheep in Iran and the Middle East 16.
Wakayama et al developed a mouse somatic cell NT method by injecting donor nuclei into enucleated oocytes using a piezo unit 6. For other researchers, obtaining results with piezo unit was difficult. Currently, numerous laboratories have reported the continual reproduction of cloned mice, including cumulus cells 6, tail-tip cells 17, Sertoli cells 18, fetal cells 19,20 and chimeric mice 21.
Cloning technology has numerous applications in industry, agriculture and medicine. This technology is considered as a research tool in basic biology 22 and is a tremendous tool for the study of processes such as nuclear reprogramming, imprinting genes and gene activation 23. The adaptability of the mouse has been introduced as an experimental model of choice, which can be used in the development of new applications 24.
Currently, progress in biological research is dependent on the capability of researchers to skillfully perform techniques in the field of stem cell and NT. In the present study, we described the successful cloning of a mouse by using piezo-assisted NT.
Materials and Methods :
Preparation of media, stock solutions and pipettes:All media, reagents and stock solutions were prepared according to a protocol by Kishigami et al 25, with the exception of mWM medium (modified Whitten's medium) which was prepared as a gift from the Department of Genetic Engineering and Animal Resources, Center for Developmental Biology, Riken Institute, Kobe, Japan.
In vitro development of fertilized embryos: evaluation of culture media and reagents: We evaluated the media quality, their supplements, and mineral oil by culturing the fertilized embryos in order to ensure optimal in vitro culture conditions. The rates of murola and blastocyst formation in mWM medium were approximately 80% (data not shown).
Animals:B6D2F1 (C57BL/6×DBA/2) mice were used to prepare oocyte and somatic-cell (cumulus cell) donors. Surrogate females were NMRI females mated with vasectomized males of the same strain. All mice were used in accordance with the Guidelines for the Care and Use of Laboratory Animals by Kurdistan University of Medical Sciences. Mice were given food and water ad libitum and maintained under controlled temperature and lighting conditions during the experiments.
Oocyte collection:Mature oocytes were collected from the oviducts of 8-12 week-old female mice. Mice were superovulated with injections of 5 IU of pregnant mare serum gonadotropin (PMSG; Serotrophin, Teikoku Hormone Mfg. Co., Tokyo, Japan) and 5 IU of human chorionic gonadotropin (hCG; Serotrophin, Teikoku Hormone Mfg. Co., Tokyo, Japan) administered 48 hr apart. We collected the oocytes 14-15 hr following the hCG injection. Oocytes were placed in M2+ hyaluronidase ((M2+Hy; Specialty Media) until the cumulus cells dispersed. The oocytes were then placed in KSOM that contained non-essential amino acids and essential amino acids (KSOM AA; Specialty Media) supplemented with 1 mg/ml bovine serum albumin (BSA, Sigma, Japan), covered with mineral oil (Sigma, Japan), and stored at 37°C, in 5% CO2 in air 25.
Preparation of donor cells:Cumulus cells were freshly isolated from recipient oocytes of B6D2F1 mice. After oocyte collection, cumulus cells were dispersed in droplets of M2 medium that contained 0.1% hyaluronidase. After 15 min, the cumulus cells were transferred to 1.5 ml tubes and centrifuged at 200 g for 5 min. The supernatant was removed and the cells suspended in M2 medium.
Enucleation of oocytes:B6D2F1 oocytes were freed from the cumulus oophorus and held in M2 medium (Specialty Media) at 37C under 5% (v/v) CO2 in air until use. The cover of a tissue culture dish was used as a micromanipulation chamber. Each ten oocytes were placed into a M2+CB (Cytochalasin B) droplet in the micromanipulation chamber and after approximately 7 min, enucleation started. Using relative contrast optics the metaphase II spindle was located inside the oocyte and identified as a translucent region. The oocyte was rotated to place the spindle between the 8 and 10 o’clock position, and then it was firmly attached to the holding pipette. The zona pellucida was "cored" following the application of several piezo-pulses (Prime Tech Ltd., Japan) to an enucleation pipette. To avoid damage to the oocyte, we maintained a large space approximately equal to the thickness of the zona pellucida that was located between the zona pellucida and the oolemma. The enucleation pipette was inserted into the oocyte without breaking the oolemma. We removed the metaphase II chromosome-spindle complex by aspiration with a minimal volume of cytoplasm. To verify enucleation, the nucleus was pushed out. It was felt solid and harder than the cytoplasm (Figure 1). The enucleated oocytes were washed three times in KSOM to remove the CB completely after which they were maintained in KSOM medium for at least 30 min in the incubator before beginning donor cell injection 25. The inner diameters of the enucleation pipettes were approximately 8 µm.
Donor nuclei injection:A very concentrated cumulus cell suspension was created in M2 medium. We picked up 3 μl of condensed donor cell suspension and introduced into a M2+PVP (Polyvinylpyrrolidone) droplet on the micromanipulation chamber. The donor cells were gently and completely mixed with M2+PVP medium using sharp forceps. The nuclei were removed from the donor cells by gentle aspiration in and out of the injection pipette until each nucleus was separated from any visible cytoplasmic material. The donor nuclei, along with a small amount of cytoplasm, were injected deep inside the ooplasm using the piezo-driven micropipette. One weak piezo pulse was applied to puncture the oolemma at the pipette tip. The donor nucleus was immediately expelled into the enucleated oocyte cytoplasm along with a minimal amount of PVP medium. The injection pipette was gently withdrawn from the oocyte. The inner diameters of the injection pipettes were 5-6 µm 25.
Oocyte activation:The injected oocytes were maintained in the M2 drop for at least 10 min, then transferred into KSOM medium and cultured for at least 30 min in an incubator at 37°C prior to activation. Next, groups of oocytes were placed in Ca2+-free CZB that contained 5 μg/ml-1 CB, 5 nM, TSA 10 mM SrCl2 (strontium chloride)) for 6 hr.
Using intact oocytes, we tested the activation medium prior to its use. Intact oocytes were maintained in activation medium for 6 hr in a 5% CO2 incubator at 37°C. During strontium treatment, the medium became turbid due to the death of approximately 5% of the oocytes.
Following activation, all resultant embryos were transferred to KSOM+TSA (5 nM, trichostatin A, Sigma) and allowed to incubate for at least 4 hr in a 5% CO2 incubator at 37°C 25.
Finally, all embryos were washed several times in mWM medium to completely remove the chemicals. For oviduct transfer, the embryos were cultured overnight until they developed to the 2-cell stage in mWM medium.
Embryo transfer:We transferred 2-cell stage cloned embryos to the oviducts of females on day 1 of pseudopregnancy (0.5 dpc). To assess development, recipients were evaluated at 19.5 days of gestation and pups were recovered from the uterus. Live pups were raised by lactation by two NMRI foster mothers.
Results :
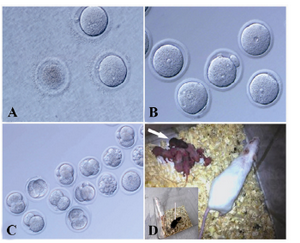
Pre- and post-implantation development of enucleated oocytes injected with cumulus cell nuclei:When we felt that we had enough experience for doing cloning experiment by NT method, we performed that seven times. Of 367 (M: 52±21) oocytes collected, 309 (85%, M: 44±14) were successfully enucleated and 299 (81%, M: 42±13) were survived. From 299 survived enucleated oocytes, 236 (81%, M: 33±9) survived following donor nuclei injection. Approximately 19% of injected oocytes died after they were transferred to activation medium. The success rate of oocyte activation was 81%. During strontium treatment, the medium became turbid due to the death of the oocytes (Figure 2). The majority of these oocytes obtained one to three pseudo-pronuclei. Of 191 oocytes, 131 (69%, M: 18±6) developed into two-cell stage embryos (Figure 2). Of these, 5 (1%, M: 0.7±1) live pups were successfully delivered, one died on the first day and two survived for two weeks. Another pup survived for one month; however the final pup survived for fourteen months. According to morphology or microanatomy of the genital system, we determined that all pups were female. We were unable to determine the color of the first mouse that died, however the remaining mice were black. The surviving cloned mouse was mated and delivered normal offspring, which she raised. Although the cloned mouse appeared obese, its offspring were of normal size.
Discussion :
In this study, we reported the production of live mouse pups by a stable NT method in which donor nuclei from cumulus cells of B6D2F1 mice were directly injected into enucleated oocytes using a piezo-actuated micromanipulator. The resultant clones derived from cumulus-cell nuclei were not contaminated, as the transferred embryos were not the result of in vitro fertilization. Pseudopregnant mice (NMRI, albino) were allowed to mate with vasectomized males (NMRI, albino) which were confirmed to be infertile. In the rare event of fertilization by a vasectomized male, the newborns would be albino. We transferred two-cell embryos into the oviducts of foster mothers. The surviving four mice had black coat colors. Where possible, assumed clones were sexed and determined to be female. Additionally, we tested all culture media and reagents, and determined their correctness.
Pioneers in the field of mouse cloning have stated that microinjections using the piezo unit, as a new technique, are of greater benefit in comparison with other micromanipulation techniques, such as ES cell injection into blastocysts or pronuclear DNA injection 25. However, personnel will need several months to achieve an adequate level of skill in the required techniques. Much practice is necessary to produce cloned mice; otherwise it is impossible 26. According to reports, when a highly skilled person performs mouse cloning experiments using cumulus cell nuclei, 99-100% of the oocytes will be enucleated successfully and 80-90% will survive nuclear injection. Of these, 70-80% should survive activation, 60-70% show pseudo-pronuclear formation and 50-60% will cleave to the two-cell stage. The rate for the production of full-term offspring ranges from 0 to 10% 25.
In the current study, with adequate technical expertise of more than one year, we could get the acceptable results in all stages of our experiment. The rate of two-cell stage development for reconstructed oocytes in the current study was higher than previous studies that used different culture media such as CZB 6,25 and KSOM 25,27. Therefore, we have shown that mWM is an appropriate medium to support the development of cloned mouse embryos.
In the current study, the donor cells were cumulus cells. There was one foster mother with five fetuses in our study. In this regard, our result is similar to the result obtained by Satoshi et al 27. They have reported that following oocyte activation, treatment of oocytes with 5-50 nM TSA increased in vitro development of somatic cloned embryos from 2- to 5-fold, which was dependent on the donor cells, including cumulus cells. The success rate of mouse cloning from cumulus cells was increased by over 5-fold without any obvious abnormalities. In their study, treatment with 5 or 50 nM TSA led to multiple conceptions. In one case, five fetuses of one foster mother were born full term. However in this study, the percent of live offspring was approximately 1%. In the study by Satoshi et al 27, it was 6%. Most likely, additional experience is necessary in order to improve the cloning experiment results in our laboratory. Since accuracy of our experiment was detected and confirmed by comparison of sex and coat color of cloned mice with source of donor cells, we didn’t perform any genetic test for more evaluation.
The adaptability of mice as a small mammal has resulted in its use as an appropriate experimental model for the study of new cloning applications and strategies 24. The information available on their reproduction, development and genetics is unmatchable to other laboratory animals 28,29. At present, numerous advanced techniques have been established by the use of mouse embryos. The mouse has a short generation period, which could be advantageous for the examining of long-term genetic effects that occur during biological manipulation, such as cloning 30,31.
Conclusion :
Our study resulted in the production of cloned mice by somatic cell NT using piezo-actuated micromanipulator. This accomplishment is another important event in the technology of cloning in Iran.
Acknowledgement :
The authors express their appreciation to Dr. Wakayama for his kind assistance. The present work was financially supported by Kurdistan University of Medical Sciences and the University of Kurdistan.
Conflict of Interest :
There is no conflict of interest in this study.
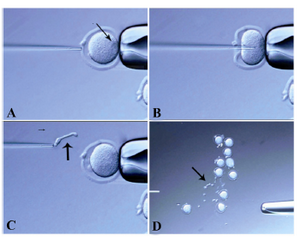
Figure 1. Oocyte enucleation. The oocyte was rotated such that the metaphase II spindle was located between the 8 and 10 o�clock positions A) Next, this oocyte was held on the holding pipette. The nucleus was removed by suction without breaking the plasma membrane. The pipette was gently pulled away from the oocyte B) and C) A group of ten oocytes follow enucleation. To verify enucleation, the nucleus was pushed out. It was felt harder than the cytoplasm D) Arrows point to nuclei
|
|
|