Orexin-1 Receptor Antagonist SB-334867 Enhances Formalin-Induced Nociceptive Behaviors in Adult Male Rats
-
 Kourosh-Arami, Masoumeh
Department of Neuroscience, School of Advanced Technologies in Medicine, Iran University of Medical Sciences, Tehran, Iran, Tel: +98 21 86701; Fax: +98 21 86701; E-mail: mkourosharami@gmail.com
Kourosh-Arami, Masoumeh
Department of Neuroscience, School of Advanced Technologies in Medicine, Iran University of Medical Sciences, Tehran, Iran, Tel: +98 21 86701; Fax: +98 21 86701; E-mail: mkourosharami@gmail.com
-
Komaki, Alireza
-
Department of Neuroscience, School of Science and Advanced Technologies in Medicine, Hamadan University of Medical Sciences, Hamadan, Iran
-
Gholami, Masoumeh
-
Department of Physiology, Medical College, Arak University of Medical Sciences, Arak, Iran
Abstract: Background: Orexin (hypocretin) is one of the hypothalamic neuropeptides that plays a critical role in some behaviors including feeding, sleep, arousal, reward processing, and drug addiction. Neurons that produce orexin are scattered mediolaterally within the Dorsomedial Hypothalamus (DMH) and the lateral hypothalamus. In the current research, we assessed the impact of prolonged application of the antagonist of Orexin Receptor 1 (OXR1) on nociceptive behaviors in adult male rats.
Methods: Sixteen Wistar rats received subcutaneous (s.c.) injections of the OXR1 antagonist, SB-334867 (20 mg/kg, i.p.), or its vehicle repetitively from Postnatal Day 1 (PND1)-PND30. On the 30th day following the final application of the OXR1 antagonist formalin-provoked pain was evaluated by injecting formalin.
Results: Administration of the OXR1 antagonist in the long-term augmented the formalin-provoked nociceptive behaviors in interphase and phase II of the formalin-induced pain.
Conclusion: Current results showed that the continued inhibiting OXR1 might be implicated in formalin-induced nociceptive behaviors. Therefore, the present study highlighted the effect of orexin on analgesia.
Introduction :
Orexin-A and -B are neuropeptides produced in the lateral hypothalamus that activate two types of receptors including Orexin Receptor 1 (OXR1) and Orexin Receptor 2 (OXR2). Orexin neuropeptides contribute to feeding, wake cycle 1,2, and reward-related processes, such as drug addiction 2-5. These peptides are produced in the Lateral Hypothalamus (LH) and activate OXR1 and OXR2 receptors, which are G-protein coupled receptors 6. OXR1 has a higher affinity to orexin-A than orexin-B, while OXR2 has the same affinity for both peptides 6. Orexin neurons are classically located in the LH, perifornical area, and dorsomedial hypothalamus. They deliver extensive afferents to several brain areas 1,6,7.
Orexins have antinociceptive effects in the brain and spinal cord in diverse forms of pain including thermal, mechanical, and chemically induced nociception. Orexin receptors have been found in several brain structures which contribute to pain processing. Orexin-A exerts analgesic impacts in the brain and spinal cord, whereas, orexin-B has shown little or no antinociceptive impacts 8. It has been shown that orexin-A displays analgesic effects analogous to morphine as measured via the hot-plate test. Some evidence displayed that the blockade of OXR1 reduced physical dependence in rats 9-11. Numerous researches have discovered that orexinergic neurons and receptors are expressed at different levels of the descending pain modulation system 12,13.
In our previous study, we showed that blockade of OXR1 results in the development of morphine dependence via reduction of cAMP response element-binding protein and phospholipase Cβ3 (PLCβ3) 14. Furthermore, PLCβ3 in the hippocampus may mediate impairment of memory by long-term blockade of OXR1 assessed by the Morris water maze 15. Moreover, chronic inhibition of OXR1 can reduce the expression of PLC in rat visual cortical neurons 16. In locus coeruleus, extracellular calcium is involved in orexin-induced postsynaptic excitation of the rat locus coeruleus neurons 17.
In the first postnatal week, just a minor subdivision of neurons in the LH is orexin-A positive 18. According to evidence, translation of orexin mRNA can be eminent at very low levels on the first postnatal day, followed by an increase to the maximum at Postnatal Day 20 (PND20) 19. Moreover, glucosensitivity 20 of LH neurons and their reaction to sensory stimuli 21 develop in PND0 to 3. Thus, orexin shows a vital role in the response of LH during development 19. Formalin injection into the rat hind paw results in agitation behavior, and this formalin-induced agitation behavior has been used as a model of inflammatory pain (the formalin test) 22. Therefore, in the current study, we want to illuminate the impact of prolonged postnatal blockade of orexin function on the nociception in rats through the formalin test.
Materials and Methods :
Animals: Wistar rats (PND1 to PND30) from the Pasteur Institute of Tehran, Tehran, Iran were located in plexiglass cages with their mothers. Temperature (22±2°C) and humidity (54±2%) were controlled. Rats were given 12-hr light/dark cycles (light at 07:00 am) and had free access to food and water. The current research was achieved based on the ethical rules of Iran University of Medical Sciences Ethics Committee, Tehran, Iran, which is based on the NIH Guide for the Care and Use of Laboratory Animals.
OXR1 antagonist, SB-334867, was dissolved in artificial Cerebrospinal Fluid (aCSF) (pH=7.4), containing 1% DMSO. The solution was divided into portions and frozen at −20°C. SB-334867 (20 mg/kg, i.p.) and its vehicle were applied every day from PND1-30 23. The rats (n=16) were divided into 2 groups: Control rats (vehicle-injected ones, n=8) and SB-injected rats (n=8).
Evaluation of nociceptive behaviors by formalin test: In the current experiment, we examined the effect of i.p. injection of SB-334867 on the formalin-evoked nociception. Formalin-induced nociception was completed in a plexiglass chamber (30×30×30 cm) with a mirror positioned beneath at a 45 angle to see the position of the animal paws. At first, the rats were adapted for 30 min in an acrylic observation chamber. 10 to 20 min after the final SB injection, formalin (50 μl; 2%) was subcutaneously injected in the plantar exterior of their right hind paw with a 25 gauge needle. Formalin administration induced a biphasic nociceptive re- sponse, including phase I (min 0-5), interphase (min 5-20), and phase II (min 20-60).
Recording of pain-related performances was initiated rapidly subsequent to injection of formalin (min 0) and sustained for 60 min. Pain-related behaviors were graded as the following; 0: the injected paw was not desirable; 1: the injected paw had little or no weight on it; 2: the due paw was raised upwards and made no contact with any surface; and 3: the paw was licked or bitten by a rat. The length of licking/biting the formalin-injected hind paw was assessed as a sign of pain-related behavior in each phase with a digital time-out stopwatch 24.
Data analysis: Data has been expressed as mean±SEM and analyzed by unpaired two-tailed Student's t-test for comparison of two groups. The defined level of statistical significance was p<0.05.
Drugs: The subsequent drugs were utilized in this study: a selective OXR1 antagonist SB-334867 (Tocris, Bristol, UK), DMSO (Sigma-Aldrich, Germany), and 2% formalin which was made by dilution of 37% formaldehyde (Temad, Tehran, Iran) via sterile saline.
Results :
Using the formalin test we explored the impact of SB-334867 on nociceptive behaviors. The pain score was assessed in each stage of the formalin-induced nociception (phase I, interphase, and II). The results revealed that long-standing injection of SB-334867 enhanced the pain-related activities made by formalin in interphase (p<0.01, Figure 1) and phase II (p<0.01, Figure 2), but not phase I (analyzed by unpaired t-test, Figure 2).
Discussion :
The present results displayed that a repeated long-lasting injection of OXR1 antagonist, SB-334867 significantly enhanced pain behaviors at inter- or late-phase of the formalin-induced pain in the present re-search. Some studies have revealed that injection of orexin-A into the periaqueductal gray may induce antinociceptive effects 25. Other evidence has demonstrated that SB-334867 can suppress stress-induced antinociception 26. Furthermore, pretreatment with orexin receptor 1 antagonist (SB 334867) decreased the antinociceptive behavioral impact of restraint stress 27.
According to the evidence, orexin may disclose an intrinsic antinociceptive result by itself. Besides, some foundations of antinociception are mediated by the orexinergic system through the activation of OXR1. Therefore, a wholly prolonged injection of SB-334867 during neural development might exhibit a vital outcome on pain, and hence cause hyperalgesia 28. Drugs of abuse induce long-lasting changes in behavior by changing synaptic function and plasticity in related brain circuits 29-32. Long-term administration of morphine, as environmental stimuli, can result in long-term alterations in behavior by changing the synaptic structure, function, and the number of synapses (the synaptic plasticity) in related brain circuits 33.
Nucleus raphe magnus as a thermoregulatory center has high concentrations of orexin receptors 7,34,35. The serotonergic neurons of raphe magnus tonically adjust nociceptive transmission 36. Thus, it may be assumed that SB-334867 may lead to hyperalgesia by altering the modulatory effect of raphe magnus on nociception. Furthermore, enhanced noxious inputs from the Nucleus Tractus Solitarius (NTS) to the Parabrachial Nucleus (PBN) after trigeminal nerve injury modulates PBN neuron activity, which accompanies the affective components of orofacial neuropathic pain 37,38.
Though SB-334867 induced hyperalgesia at the inter- or late-phase of formalin-induced pain, it failed to affect nociception in phase I. The transient early phase I, which is acute pain, imitates the activation of nociceptive sensory C-fiber {Martindale, 2001 #102}by formalin 29,32. Hence, while persistent administration of SB-334867 did not change nociceptor transduction and transformation through afferent C fibers of pain, it could modify the inter- or late-phase of the formalin-induced pain.
The formalin injection in the paw causes biphasic flinching of the injected paw. Intrathecal application of orexin-A reduced the summation of flinches in phases 1 and 2 in the formalin test. These impacts of orexin-A were entirely antagonized by pre-treatment with SB-334867, a selective orexin-1 receptor antagonist. Furthermore, intrathecal injection of orexin-A blocked the expression of Fos-like immunoreactivity 39, caused by paw formalin injection, in laminae I-II of L4-5 of the spinal cord. Therefore, the spinal orexin-1 receptor is contributed to the nociceptive transmission and the activation of the spinal orexin-1 receptor results in analgesic effects in the rat formalin test 22. In addition, intracerebroventricular application of orexin-A has been reported to induce an antinociceptive impact in the rat hot plate test 40.
Conclusion :
In conclusion, according to the current results, it seems that orexin might act on the signaling pathways associated with nociception to provide a novel possible therapeutic goal in pain treatment. Nonetheless, additional in vivo and in vitro research are required to clarify the role of the orexinergic system in the modulation of pain.
Acknowledgement :
This work was funded by Iran University of Medical Sciences (code: 30368).
Funding: The authors thank Iran University of Medical Sciences for financial support.
Conflict of Interest :
The authors have no financial or proprietary interests in any material discussed in this article.
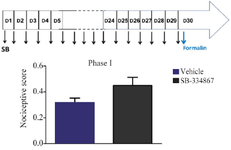
Figure 1. The effect of long-term infusion of SB-334867 on formalin-induced pain-related behaviors. Upper schematic plan demonstrates the experimental protocols used for assessment of nociceptive behaviors in MK-treated rats. Bar chart for chronic injection of SB-334867 in the formalin-induced pain displays mean nociceptive score of phase 1 (min 0–5) compared to Vehicle group. Recording of the nociceptive behaviors started instantly for 60 min following formalin injection (2% in saline, 50 µl, s.c.) into the hind paw (minute 0). Data is expressed as mean ± SEM. n=8 per group.
|

Figure 2. The persistent injection of SB-334867 on formalin-produced nociception. The graph shows the impact of chronic injection of SB-334867 on the formalin-induced pain and displays mean nociceptive score of interphase and phase 2 compared to Vehicle group. Measuring the nociceptive behaviors initiated instantly for 60 minutes after formalin injection (2% in saline, 50 µl, s.c.) into the hind paw (min 0). Data is presented as mean ± SEM. *p<0.01 compared to SB-vehicle (Vehicle), n=8 per group.
|
|