Association of ATP-Binding Cassette Transporter A1 (ABCA1)-565 C/T Gene Polymorphism with Hypoalphalipoproteinemia and Serum Lipids, IL-6 and CRP Levels
-
 Halalkhor, Sohrab
Department of Biochemistry, Faculty of Medicine, Babol University of Medical Sciences, Babol, Iran, Tel: +98 21 22432020, E-mail: sohrabhalalkhor@yahoo.com
Halalkhor, Sohrab
Department of Biochemistry, Faculty of Medicine, Babol University of Medical Sciences, Babol, Iran, Tel: +98 21 22432020, E-mail: sohrabhalalkhor@yahoo.com
-
Department of Biochemistry, Faculty of Medicine, Babol University of Medical Sciences, Babol, Iran
-
Moradi Firouzjah, Hamid
-
Department of Molecular and Cell biology, Faculty of Basic Sciences, University of Mazandaran, Babolsar, Iran
-
Parsian, Hadi
-
Department of Biochemistry, Faculty of Medicine, Babol University of Medical Sciences, Babol, Iran
-
Jalali, Seyed Farzad
-
Department of Cardiology, Faculty of Medicine, Babol University of Medical Sciences, Babol, Iran
-
Babashamsi, Mohammad
Monoclonal Antibody Research Center, Avicenna Research Institute, ACECR, Tehran, Iran, Tel: +98 21 22432020, E-mail: babashams@avicenna.ac.ir
-
Monoclonal Antibody Research Center, Avicenna Research Institute, ACECR, Tehran, Iran
Abstract: Background: ATP-binding cassette transporter A1 (ABCA1) is a membrane integral protein which plays a vital role in High Density Lipoprotein (HDL) metabolism and exerts a protective effect against Hypoalphalipoproteinemia (HA) by mediation of rate-limiting step in HDL biogenesis. In addition, this protein possesses anti-inflammatory effects by inhibiting the production of some inflammatory cytokines in macrophages. This study investigated the association of ABCA1-565 C/T gene polymorphism with HA and serum lipids, IL-6 and CRP levels.
Methods: A population which consisted of 101 HA and 95 normal subjects were genotyped for ABCA1-565C/T polymorphism by Polymerase Chain Reaction-Restriction Fragment Length Polymorphism (PCR-RFLP). The serum concentrations of lipids, IL-6 and high sensitive-CRP (hs-CRP) were measured by the relevant methods.
Results: The frequency of T allele was significantly higher in the HA group than the controls (31.7 vs. 19.5%, p=0.002). Thus, carriers of the T allele (CT and TT genotypes) had a higher risk for HA (p=0.016, OR=2.04, 95% CI=1.14-3.63). T allele carriers demonstrated decreased HDL-C and increased triglyceride, IL-6 and CRP levels than those with the CC genotype.
Conclusion: This study suggests that the-565 C/T polymorphism of ABCA1 gene is associated with an increased risk of HA, decreased HDL-C and increased TG, IL-6 and CRP.
Introduction :
Hypoalphalipoproteinemia (HA) is a dyslipidemia which is characterized by HDL-C concentration below 1.04 mmol/L (40 mg/dL) 1. High Density Lipoprotein (HDL) is a multifunctional lipoprotein particle which plays a vital role in the metabolism of lipids and protection against a number of diseases 2,3 by I) removing excess cholesterol from tissues and transporting them to liver for elimination; II) protecting the endothelial cell function; III) inhibiting the LDL oxidation and platelet aggregation and IV) participating in lipid metabolism by transferring the lipase cofactor and receptor. HA is associated with an increased risk of Cardiovascular Disease (CVD), type 2 diabetes mellitus, dementia, lung cancer and lymphoma 4,5. Its occurrence may be due to environmental factors and lifestyle (e.g. smoking, medications, overweight, physical inactivity) as well as genetic defects 6.
The most severe form of HA is the Tangier Disease (TD), a disorder which is characterized by near zero level of HDL. Homozygosity for mutations in ABCA1 gene leads to TD 3. ABCA1 gene is located in the q31 region of chromosome 9 and consists of 50 exons that encode a 2261 amino acid integral membrane protein 7. ABCA1 mediates the rate-limiting step in HDL biogenesis by transporting cellular excess free cholesterol and phospholipids to an apolipoprotein acceptor 8. It is broadly expressed in macrophages, hepatocytes, intestinal cells, lung cells, adrenal gland and placental trophoblast 9. This expression is highly induced by sterols, through nuclear Liver X Receptor (LXR)/Retinoid X Receptor (RXR) pathway 10.
The accumulation of sterol-rich lipoproteins, together with their oxidation in the artery wall, results in the recruitment of macrophages and the production of inflammatory molecules 11. A study by Tang et al indicated that the macrophage ABCA1, in addition to cholesterol efflux, functioned as an anti-inflammatory receptor. The interaction of apolipoprotein (apo) A-I with ABCA1-expressing cells stimulates autophosphorylation (activation) of Janus kinase2 (JAK2) by an ABCA1-dependent mechanism. Activated JAK2 has two independent effects: I) it enhances the apo A-I binding activity of ABCA1 which is essential for lipid export; II) Phosphorylation and activation of Signal Transducer and Activator of Transcription3 (STAT3). The activated STAT3 has anti-inflammatory function in macrophages. The phosphorylated STAT3 migrates to the nucleus and regulates the transcription of proteins which repress the production of inflammatory cytokines IL-1β, IL-6, and TNF-α 12. Regarding the increase of the mentioned inflammatory cytokines, especially IL-6 and to a lesser degree TNF-α and IL-1β, the liver and aortic endothelial cells produces C-Reactive Protein (CRP) 13.
Although the TD is rare, common Single Nucleotide Polymorphisms (SNPs) in the ABCA1 gene are present in the general population 14. Consequently, much attention has been paid to the relationship between ABCA1 gene polymorphisms with different phenotypes, including lipid variables and clinical endpoints 15,16. One of the common SNPs is -565C/T (rs2422493), which serves as a cytosine to thymine substitution in the promoter region of the ABCA1 gene, which affects expression, resulting in the formation of promoter low-activity (C/T, T/T) and high-activity (C/C) genotypes 17. The previous association studies on this polymorphism were limited to various forms of CVD, as well as changes in the lipid profile. Of course, their results were inconsistent 7,18.
As HA is the most common dyslipidemia in Iranian population 19, and moreover there exists no data yet on the association of the ABCA1 gene polymorphisms with serum inflammatory markers, the present study investigated the association of ABCA1 -565 C/T polymorphism with HA and serum lipids, IL-6 and CRP levels in an Iranian population.
Materials and Methods :
Subjects: This research was approved by the Ethics Committee of Babol University of Medical Sciences and the researches undertook Helsinki treaty. Participants in the study were selected among a large number of individuals who referred to the laboratories of Babol city hospitals from January 2013 to August 2014. HA subjects were selected among the individuals with HDL-C <40 mg/dL and the control group among the individuals with HDL-C ≥40 mg/dL who were accepted to participate in the study. Written, informed consent was obtained from the volunteers. Their personal data including age, residence, past medical history, drugs, cigarette and alcohol consumption were collected through an interview. Anthropometric characteristics of the included individuals were measured and the Body Mass Index (BMI) was calculated for each individual (weight in kilograms/height in meters squared). Individuals with a BMI> 30, consumers of cigarettes, alcohol and those with diabetes, rheumatic fever, tuberculosis, infectious diseases accompanied by fever, diseases of liver and thyroid, pregnants and patients treated with lipid-lowering agents, anti-inflammatory and anti-cancer drugs were excluded from the study. The remaining 343 subjects were referred to the Cardiology department of Rouhani Hospital (Babol, Iran). Eventually, 196 subjects enrolled in the study and based on HDL-C levels, were divided into two groups: HA (HDL-C<40 mg/dL) and normal (HDL-C≥40 mg/dL).
Sample collection: 3 ml of blood sample was taken from each subject after overnight fasting. 1.5 ml of blood was transferred to a tube containing Ethylene Diamine Tetra Acetic acid (EDTA) for DNA extraction and 1.5 ml was transferred to a tube without any anticoagulant agent to obtain serum for biochemical analysis.
Biochemical analysis
Lipid profile analysis: The serum total cholesterol, LDL-cholesterol (LDL-C) and triglycerides were measured by enzymatic colorimetric methods. HDL-C was determined after precipitation of apoB-containing lipoproteins with dextran sulfate and magnesium. All measurements were done using Pars Azmun diagnostic kits (Pars Azmun, Iran).
Inflammatory factors analysis: CRP was determined by turbidimetric immunoassay method with auto-analyzer (Hitachi 902, Japan), using hs-CRP diagnostic kit (Pars Azmun, Iran). IL-6 level was determined using enzyme-linked immunosorbent assay (ELISA) method (Pelikine Compact human IL-6 ELISA kit, CLB, Amsterdam, the Netherlands).
Genotyping of ABCA1 -565 C/T polymorphism analysis: Genomic DNA was extracted from peripheral blood leukocytes using salting out method 20. Genotyping was accomplished using PCR-RFLP technique. For amplification of fragment containing -565 C/T polymorphism, a pair of forward primer 5/-CTCGGG TCCTCTGAGGGACCT-3/and reverse primer 5/-CC GCAGACTCTCTAGTCCAC-3/(CinnaGen Co, Iran) was used. PCR reaction mixture, having a final volume of 50 μl, contained 2 μl DNA, 2 μl (10 μM) of each primers, 4 μl (2.5 mM) of dNTPs, 3 μl (25 mM) of MgCl2, 5 μl of 10x buffer, 2.5 U of Taq DNA polymerase (Fermentas, Burlington, USA) and the remaining was DDW. The reaction mixture was denatured at 95°C for 2 min, followed by 35 cycles of denaturation at 94°C for 30 s, annealing at 60°C for 60 s and extension at 72°C for 60 s, with a final extension at 72°C for 10 min. After amplification, aliquots of the PCR products were digested with 2U AciI restriction enzyme (Thermo scientific, USA) at 37°C overnight. Separation of the fragments obtained after digestion was done using 12% Polyacrylamide Gel Electrophoresis (PAGE) for 3 hr, under constant voltage (200 V). The gel was stained with silver nitrate and the bands were visualized. The AciI restriction enzyme has the ability to recognize the (C/CGC) sites. After digestion of 351 bp fragment, three possible genotypes were distinguished: homozygous CC (148, 130 and 73 bp), heterozygous CT (278, 148, 130 and 73 bp), and homozygous TT (278 and 73 bp).
Statistical analysis: Data were analyzed statistically using the SPSS 18 software (SPSS Inc., Chicago, IL, USA), and were expressed as mean±standard deviation (M±SD). hs-CRP and IL-6 concentrations were natural log transformed to improve normality. An independent T-test was used for comparison of the demographic and biochemical variables between the two groups of HA and control. Allele frequencies were calculated by the gene counting method and differences in the frequency distributions of the alleles and genotypes between HA and control groups were evaluated by Chi-square (χ2) test. Hardy-Weinberg equilibrium (HWE) for both of the cases and control groups was assessed by the χ2 test. Multivariate logistic regression was performed to analyze the relationship between -565 C/T polymorphism and HA that were adjusted for covariates (age, BMI and sex). The biochemical parameters evaluated based on genotypes using multivariate analysis of variance (ANOVA) that were adjusted for covariates as mentioned above. The p-value less than 0.05 was considered statistically significant.
Results :
Table 1 presents the demographic and biochemical data of HA and control groups. There existed a significant difference in total cholesterol, triglycerides, HDL-C, LDL-C, IL-6 and hs-CRP between the two groups (p<0.05), however, there were no significant differences between the two groups in sex, age and BMI (p>0.05).
The frequency distribution of the alleles and genotypes in ABCA1-565 C/T polymorphism between HA and control groups, are shown in table 2.
There was consistency in the frequency distribution of -565 C/T polymorphism in the HA and control groups with the HWE. It also showed that the frequency of T allele was significantly higher among the HA group in comparison with the controls. In addition, the results illustrated that the distribution of genotypes between the two groups was significantly different.
A multivariate logistic regression was performed to evaluate the association of -565 C/T polymorphism with HA. When the CC genotype was used as a reference, the TT and CT genotypes were both significantly associated with HA risk. So that, a significant association was observed for T allele carrier genotypes when compared with CC genotype for HA risk (Table 3).
The analysis of biochemical parameters, based on genotypes, demonstrated that the T allele carriers exhibited significantly lower HDL-C levels and significantly higher TG, IL-6 and hs-CRP levels than those with the CC genotype (p<0.05), while the total cholesterol and LDL-C were not significantly different among genotypes (p>0.05, Table 4).
Discussion :
The results of the current study showed an association between -565 C/T polymorphism and the risk of HA. Based on the results, individuals carrying the T allele were more susceptible to HA.
Previous studies investigated the effect of -565 C/T polymorphism on HA-related diseases, such as various types of CVD. The results of these studies have been controversial. Lutacuta et al showed that the presence of the T allele in -565 C/T locus was associated with the severity of the atherosclerosis in patients with established Coronary Artery Disease (CAD) 18. Benton et al in a Multi-Ethnic Study of Atherosclerosis (MESA) reported a borderline association between the presence of T allele and higher prevalence of the coronary artery calcification 14. In contrast, Jensen et al reported an inverse relationship between the -565 C/T variant with the risk of Coronary Heart Disease (CHD) among healthy women 21.
Of course, there existed no association in a number of studies. For example, in a study of 316 Tunisian patients who underwent coronary angiography, according to Rejeb et al there was no significant association between -565 C/T polymorphism with the stenosis risk 7. In addition, Takagi et al found no relationship between -565 C/T with myocardial infarction or angina pectoris in Japanese patients 22.
The reason for these conflicting results obtained in various studies is still unclear. Racial difference of different populations might be one of the major reasons for disputes. In addition, the presence or absence of an observed association in any race, ethnicity or geographic populations may be related to a number of other factors, such as gene-gene interactions and environmental factors.
Also, there have been contradictory results regarding the association between -565 C/T variant and lipid profile. While some investigators did not find any significant association between the polymorphism and lipid profile 7,14,18, Liu et al found a significant decrease of HDL-C concentrations in TT genotype compared to those with the CC genotype 23. In the present study, the assessment of lipid profile based on genotype showed that T allele carriers had significantly lower HDL-C and higher TG levels than those with the CC genotype. In addition, this study evaluated the association of -565 C/T polymorphism with IL-6 and CRP levels for the first time. The results revealed the association between this polymorphism, with a significant increase of IL-6 and CRP levels. The -565 C/T polymorphism is located in the promoter region. Hence, a possible mechanism for the observed associations may be due to the effect of -565 C/T polymorphism on the ABCA1 gene expression level. Studies of Kyriakou et al on the ABCA1 expression levels in ex vivo atherosclerotic tissue of individuals with different genotypes of -565 C/T exhibited a lower expression level in T allele carriers. In addition, in vitro studies in macrophages revealed remarkable results. Transient transfection and reporter assays demonstrated a lower activity of the T allelic promoter in driving gene expression than the C allelic promoter. Also, investigation of allele specific effect of the -565 C/T polymorphism on binding of nuclear proteins to the ABCA1 gene promoter indicated that this variant was located in a transcription factor binding site and lower binding at the presence of the T allele 17. These data suggest that the polymorphism -565 C/T may have effect on the expression of ABCA1 and result in a reduction in the transportation of cholesterol and anti-inflammatory activity; resulting in decreased HDL-C and increased IL-6, and consequently CRP.
The increase of TG in T allele carriers of the -565 C/T variant is remarkable. The activity of Cholesterol Ester Transfer Protein (CETP) causes the equilibration of the core components of lipoprotein particles. Cholesteryl esters are transferred from HDL to TG-rich lipoproteins, while TG is transferred by TG-rich lipoproteins to HDL. Decreased ABCA1 activity, resulting in decreased HDL-C, may decrease the cholesteryl ester/TG exchange. TG component of HDL is hydrolyzed by hepatic lipase. Thus, decreased transfer of TG to HDL may ultimately decrease the TG catabolism 24. In addition, studies on ABCA1 deficient mice suggested the role of hepatocyte ABCA1 in TG metabolism. Based on these results, large nascent HDL particles assembled by hepatocyte ABCA1 bind to a putative membrane receptor (Topβ, target of pre-β), which in turn stimulates PI3 kinase activation and as a result reduces the lipid mobilization and secretion of normal-sized VLDL particles (VLDL2). In the absence of hepatic ABCA1 or diminished ABCA1 activity, reduced nascent HDL particle formation leads to diminished signaling through Topβ, which results in a reduction of PI3 kinase activation, increased lipid mobilization, and increased secretion of larger TG-enriched VLDL particles (VLDL1) 25,26. Thus, it is possible that the impact of -565 C/T polymorphism on HDL-C levels results to an increase in the TG level.
Conclusion :
In conclusion, this study suggests that the ABCA1-565 C/T gene polymorphism is associated with an increased risk of HA, decreased HDL-C and increased TG, IL-6and CRP. Due to the association of HA with a number of diseases, the identification of genetic risk factors associated with HA is an opportunity to identify individuals that are susceptible to this disorder, and also for pre-emptive measures to be taken in order to prevent HA, and consequently the related diseases. On the other hand, an evaluation of the ABCA1 variants association with the inflammatory factors can lead to the recognition of the agent involved in progression of inflammation. Hence, further study with larger sample sizes is recommended in order to confirm the findings of this study.
Acknowledgement :
The authors are grateful to Babol University of Medical Sciences for financial support (Grant No: 2504), Dr. Kourosh Kamali and Mr. Kioomars Saliminejad, Reproductive Biotechnology Research center, Avicenna Research Institute for scientific co-operations.
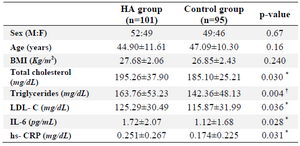
Table 1. The demographic and biochemical data of the study population
n: number of individuals
The results are shown as mean±SD; *p<0.05, †p<0.01.
|
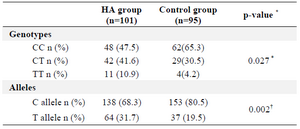
Table 2. Genotype and allele frequencies of the -565 C/T gene polymorphism in HA and control group
n: number of individuals; *p<0.05, †p<0.01
a: χ2-test for distributions of genotype and allele frequencies between the HA and control group.
|
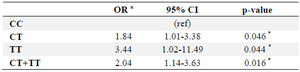
Table 3. Association of ABCA1-565 C/T polymorphism with HA
a: Adjusted ORs were obtained from a multivariate logistic regression with
adjustment for age, sex and BMI.
* p<0.05.
|
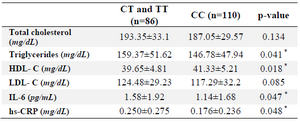
Table 4. The levels of biochemical parameters according to the genotype
The differences between means were analyzed by multivariate ANOVA adjusted for age, sex and BMI.
* p<0.05.
|
|